Sinir sistemlerinin evrimi
Sinir sistemlerinin evrimi, hayvanlarda (veya metazoanlarda) sinir sistemlerinin ilk gelişimine kadar uzanır. Nöronlar, hareketli tek hücreli ve kolonyal ökaryotlarda bulunan aksiyon potansiyellerinin mekanizmasını uyarlayarak çok hücreli hayvanlarda özel elektrik sinyal hücreleri olarak geliştirildi. Karmaşık protozoalarda bulunanlar gibi birçok ilkel sistem, hareketlilik ve hayatta kalmak için gerekli diğer yönler için elektriksel olmayan sinyalleme kullanır. Veriler, mesajlaşma için kimyasal bir gradyan kullanan bu sistemlerin bugün bilinen elektrik sinyal hücrelerine dönüştüğünü gösteriyor.[1]
İlk olarak Cnidaria (denizanası) gibi hayvanlarda görülen basit sinir ağları, motor ve duyusal işlevlerde ikili bir amaca hizmet eden polimodal nöronlardan oluşuyordu. Cnidarianlar, her ikisi de denizanası olmasına rağmen çok farklı sinir sistemlerine sahip olan Ctenophores (tarak denizanası) ile karşılaştırılabilir. Cnidarians'tan farklı olarak, Ctenophores, elektrokimyasal sinyalleme kullanan nöronlara sahiptir. Bu şaşırtıcıydı çünkü Ctenophora şubesinin, sinir sistemi olmayan Porifera'dan (süngerler) daha eski olduğu düşünülüyordu.
Bu, erken sinir sisteminin nasıl ortaya çıktığını açıklayan iki teorinin ortaya çıkmasına yol açtı. Bir teori, sinir sisteminin tüm bu şubelerin temel bir atasında ortaya çıktığını, ancak Porifera'da kaybolduğunu belirtti. Diğer teori, sinir sisteminin bağımsız olarak iki kez (birlikte evrim), biri bazal Cnidarianlar ve bazal taraklılar için ortaya çıktığını belirtir.
İki taraflı hayvanlar - omurgasızlarda ventral sinir kordonları ve kordalılarda bir notokord tarafından desteklenen dorsal sinir kordonları - merkezi bir bölge etrafında bulunan bir merkezi sinir sistemi ile evrimleşmiştir, bu süreç sefalizasyon olarak bilinir.
Sinir öncüleri
Nöral aktivite için gerekli olan aksiyon potansiyelleri tek hücreli ökaryotlarda gelişmiştir. Bunlar sodyum aksiyon potansiyelleri yerine kalsiyum kullanır, ancak mekanizma muhtemelen çok hücreli hayvanlarda nöral elektrik sinyalleşmesine uyarlanmıştır. Obelia gibi bazı sömürge ökaryotlarında, elektrik sinyalleri yalnızca sinir ağları aracılığıyla değil, aynı zamanda koloninin ortak sindirim sistemindeki epitel hücreleri aracılığıyla da yayılır. [2] Koanoflagellatlar, filasterea ve mesomycetozoea dahil olmak üzere birçok metazoan olmayan filumun, salgı SNARE'leri, Shank ve Homer dahil olmak üzere sinaptik protein homologlarına sahip olduğu bulunmuştur. Koanoflagellatlarda ve mesomycetozoea'da, bu proteinler kolonyal evrelerde yukarı doğru düzenlenir ve bu proto-sinaptik proteinlerin hücreden hücreye iletişim için önemini ortaya koyar.[3] Evrimde nöronların ve ilk sinir sistemlerinin nasıl ortaya çıktığına dair fikirlerin tarihi, Michel Antcil tarafından 2015 yılında yayınlanan bir kitapta tartışıldı.[4]
Süngerler
Süngerlerin birbirine sinaptik bağlantılarla bağlı hücreleri, yani nöronları ve dolayısıyla sinir sistemi yoktur. Bununla birlikte, sinaptik işlevde kilit rol oynayan birçok genin homologlarına sahiptirler. Son zamanlarda yapılan araştırmalar, sünger hücrelerinin, bir postsinaptik yoğunluğa (sinapsın sinyal alan kısmı) benzeyen bir yapı oluşturmak üzere bir araya toplanmış bir grup proteini ifade ettiğini göstermiştir. Ancak, bu yapının işlevi şu anda belirsizdir. Sünger hücreleri sinaptik iletim göstermese de, birbirleriyle kalsiyum dalgaları ve tüm vücudun kasılması gibi bazı basit eylemlere aracılık eden diğer uyarılar yoluyla iletişim kurarlar. Sünger hücrelerin komşu hücrelerle iletişim kurmasının diğer yolları, hücre zarlarının oldukça yoğun bölgeleri boyunca veziküller taşımadır. Bu veziküller iyonları ve diğer sinyal moleküllerini taşır, ancak gerçek bir sinaptik işlev içermez.[5]
Sinir ağları

Denizanası, taraklılar ve ilgili hayvanlar, merkezi bir sinir sisteminden ziyade yaygın sinir ağlarına sahiptir. Çoğu denizanasında sinir ağı vücuda az çok eşit olarak yayılır; tarak jölelerinde ağzın yakınında konsantre edilir. Sinir ağları, kimyasal, dokunsal ve görsel sinyalleri alan duyusal nöronlardan, vücut duvarının kasılmalarını aktive edebilen motor nöronlardan ve duyusal nöronlardaki aktivite modellerini algılayan ve motor nöron gruplarına sinyaller gönderen ara nöronlardan oluşur. sonuç. Bazı durumlarda, ara nöron grupları ayrı gangliyonlar halinde kümelenir.[6]
Işınsal simetrili canlılarda, sinir sisteminin gelişimi nispeten yapılandırılmamıştır. Bilaterilerin aksine, radiata sadece iki ilkel hücre katmanına sahiptir, endoderm ve ektoderm . Nöronlar, diğer tüm ektodermal hücre tipleri için öncü görevi gören özel bir ektodermal öncü hücreler grubundan üretilir.[7]
Sinir kordonları

Mevcut hayvanların büyük çoğunluğu iki taraflıdır, yani birbirlerinin yaklaşık ayna görüntüsü olan sol ve sağ tarafları olan hayvanlar. Tüm bilaterilerin, 550-600 milyon yıl önce Ediakara döneminde ortaya çıkan solucan benzeri ortak bir atadan geldiği düşünülmektedir. Temel iki taraflı vücut formu, ağızdan anüse uzanan içi boş bir bağırsak boşluğuna sahip bir tüp ve ön tarafında özellikle büyük bir ganglion bulunan bir sinir kordonudur, buna "beyin" denir.
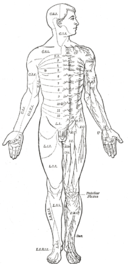
İnsanlar da dahil olmak üzere memeliler dahi, sinir sistemi düzeyinde parçalı çift taraflı vücut planını gösterir. Omurilik, her biri vücut yüzeyinin bir kısmını innerve eden motor ve duyusal sinirlere yol açan bir dizi segmental gangliyon içerir ve alttaki kaslar. Ekstremitelerde innervasyon paterninin düzeni karmaşıktır, ancak gövdede bir dizi dar banda yol açar. İlk üç bölüm beyne aittir ve ön beyin, orta beyin ve arka beyine yol açar.[8]
Bilateriyenler, embriyonik gelişimin çok erken döneminde meydana gelen olaylara dayanarak, protostomlar ve deuterostomlar olarak adlandırılan iki gruba (üst şube) ayrılabilir. Deuterostomlar omurgalıların yanı sıra derisidikenlileri ve hemikordatları (esas olarak meşe palamudu solucanları) içerir. Daha çeşitli grup olan protostomlar, eklembacaklıları, yumuşakçaları ve sayısız solucan türünü içerir. Sinir sisteminin vücuttaki yerleşiminde iki grup arasında temel bir fark vardır: protostomlarda vücudun ventral (genellikle alt) tarafında bir sinir kordonu bulunurken, deuterostomlarda sinir kordonu dorsalde (genellikle üstte) bulunur.) yan. Aslında, dorsalden ventral gradyanlar gösteren birkaç genin ekspresyon paternleri de dahil olmak üzere, vücudun birçok yönü iki grup arasında ters çevrilmiştir. Bazı anatomistler şimdi, protostom ve deuterostom gövdelerinin birbirlerine göre "ters çevrildiğini" düşünüyorlar; bu, ilk olarak Geoffroy Saint-Hilaire tarafından böcekler için omurgalılara kıyasla önerilen bir hipotezdir. Bu nedenle, örneğin böcekler vücudun karın orta hattı boyunca uzanan sinir kordonlarına sahipken, tüm omurgalıların sırt orta hattı boyunca uzanan omurilikleri vardır.
Annelida
Solucanlar, vücudun uzunluğu boyunca uzanan ve kuyruk ve ağızda birleşen çift sinir kordonlarına sahiptir. Bu sinir kordonları, bir merdivenin basamakları gibi enine sinirlerle birbirine bağlanır. Bu enine sinirler, hayvanın iki tarafını koordine etmeye yardımcı olur. Baş ucundaki iki ganglion, basit bir beyne benzer şekilde çalışır. Hayvanın göz noktalarındaki fotoreseptörler, aydınlık ve karanlık hakkında duyusal bilgi sağlar.[9]
Nematoda
Çok küçük bir solucanın sinir sistemi, yuvarlak solucan Caenorhabditis elegans, sinaptik seviyeye kadar haritalandırılmıştır. Her nöron ve onun hücresel soyu kaydedilmiştir ve hepsi olmasa da çoğu nöral bağlantı bilinmektedir. Bu türde sinir sistemi cinsel olarak dimorfiktir; iki cinsiyetin, erkeklerin ve hermafroditlerin sinir sistemleri, cinsiyete özgü işlevleri yerine getiren farklı sayıda nörona ve nöron gruplarına sahiptir. C. elegans'ta erkeklerin tam olarak 383 nöronu varken, hermafroditlerin tam olarak 302 nöronu vardır.
Eklembacaklılar

Eklembacaklılar gibi böcekler ve kabuklular, sinir sisteminin bir dizi yapılmış olması gangliyon ile bağlanmış, ventral sinir kablosunun uzunluğu boyunca uzanan iki paralel eklemlerin oluşan göbek .[10] Tipik olarak, her vücut segmentinin her iki tarafında bir ganglion bulunur, ancak bazı ganglionlar beyni ve diğer büyük gangliyonları oluşturmak için kaynaşmıştır. Baş segmenti, supraözofageal ganglion olarak da bilinen beyni içerir. Böcek sinir sisteminde beyin anatomik olarak protoserebrum, deutoserebrum ve tritoserebrum'a bölünmüştür . Beynin hemen arkasında, üç çift kaynaşmış gangliyondan oluşan subözofageal ganglion bulunur. Ağız kısımlarını, tükürük bezlerini ve bazı kasları kontrol eder . Birçok eklembacaklı, görme için bileşik gözler ve koku alma ve feromon hissi için antenler dahil olmak üzere iyi gelişmiş duyu organlarına sahiptir. Bu organlardan gelen duyusal bilgiler beyin tarafından işlenir.
Böceklerde birçok nöron, beynin kenarında konumlanmış ve elektriksel olarak pasif olan hücre gövdelerine sahiptir - hücre gövdeleri yalnızca metabolik destek sağlamaya hizmet eder ve sinyalleşmeye katılmaz. Protoplazmik bir lif hücre gövdesinden çıkar ve bolca dallanır, bazı kısımları sinyalleri iletir ve diğer kısımları sinyalleri alır. Bu nedenle, böcek beyninin çoğu parçası, çevre çevresinde düzenlenmiş pasif hücre gövdelerine sahipken, nöral sinyal işleme, iç kısımda, nöropil adı verilen bir protoplazmik lifler yumağı içinde gerçekleşir.[11]
Merkezi sinir sistemlerinin evrimi
İnsan beyninin evrimi
Modern insanın ataları, insanın evrim zaman çizelgesi boyunca (bkz. İnsanın evrimi) ilerledikçe, Homo habilis'te yaklaşık 600 cm³'ten Homo neanderthalensis'te 1736 cm³'e kadar, beyin hacminde kademeli bir artış olmuştur. Bu nedenle, genel olarak beyin hacmi ile zeka arasında bir ilişki vardır.[12] Bununla birlikte, modern Homo sapiens, neandertallerden daha küçük bir beyin hacmine (beyin boyutu 1250 cm³) sahiptir; kadınların beyin hacmi erkeklerden biraz daha küçüktür ve "hobbit" lakaplı Flores hominidlerinin (Homo floresiensis) kafatası kapasitesi yaklaşık 380 cm³, yani Homo erectus ortalamasının yaklaşık üçte biri kadardı ve bir şempanze için küçük kabul ediliyordu. Bir insular cücelik vakası olarak H. erectus'tan evrimleştikleri ileri sürülmektedir. Üç kat daha küçük beyinlerine rağmen, H. floresiensis'in ateş kullandığına ve önerilen ataları H. erectus kadar sofistike taş aletler yaptığına dair kanıtlar var.[13] Iain Davidson, insan beyni boyutundaki zıt evrimsel kısıtlamaları "İhtiyacınız olduğu kadar büyük ve olabildiğince küçük" olarak özetler.[14] İnsan beyni, türün varlığı boyunca uğraştığı metabolik, çevresel ve sosyal ihtiyaçlar etrafında gelişmiştir. İnsansı türler artan beyin boyutu ve işlem gücü ile evrimleştikçe, genel metabolik ihtiyaç arttı. Şempanzelerle karşılaştırıldığında, insanlar hayvanlardan bitkilerden daha fazla kalori tüketir. Kesin olmamakla birlikte, araştırmalar, diyetteki bu değişimin, hayvansal ürünlerde daha kolay bulunan yağ asitlerine artan ihtiyaçtan kaynaklandığını göstermiştir. Bu yağ asitleri beyin bakımı ve gelişimi için gereklidir. Göz önünde bulundurulması gereken diğer faktörler, sosyal etkileşim ihtiyacı ve hominidlerin zaman içinde çevreleriyle nasıl etkileşime girdiğidir.[15]
Beyin evrimi, paleonöroloji adı verilen bir nöroloji ve paleontoloji dalı olan endokastlar kullanılarak incelenebilir.
Ayrıca bakınız
Kaynakça
- ^ "nervous system | Definition, Function, Structure, & Facts". Encyclopedia Britannica (İngilizce). 13 Haziran 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Nisan 2021.
- ^ "Evolution of nervous systems". Neurobiology: molecules, cells, and systems. Wiley-Blackwell. 2001. s. 21. ISBN 978-0-632-04496-2.
- ^ Burkhardt (1 Eylül 2017). "Evolutionary origin of synapses and neurons - Bridging the gap". BioEssays. 39 (10): 1700024. doi:10.1002/bies.201700024. ISSN 0265-9247. PMID 28863228.
- ^ Dawn of the Neuron: The Early Struggles to Trace the Origin of Nervous Systems. Montreal & Kingston, London, Chicago: McGill-Queen's University Press. 2015. ISBN 978-0-7735-4571-7.
- ^ Leys (15 Şubat 2015). "Elements of a 'nervous system' in sponges". Journal of Experimental Biology (İngilizce). 218 (4): 581-591. doi:10.1242/jeb.110817. ISSN 0022-0949. PMID 25696821. 20 Kasım 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 26 Haziran 2021.
- ^ Ruppert, Edward E.; Fox, Richard S.; Barnes, Robert D. (2004). Invertebrate zoology : a functional evolutionary approach. Internet Archive. Belmont, CA : Thomson-Brooks/Cole. ISBN 978-0-03-025982-1.
- ^ Development of the nervous system. Academic Press. 2006. ss. 3-4. ISBN 978-0-12-618621-5.
- ^ Ghysen A (2003). "The origin and evolution of the nervous system". Int. J. Dev. Biol. 47 (7–8): 555-62. PMID 14756331. 19 Haziran 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 26 Haziran 2021.
- ^ ADEY WR (February 1951). "The nervous system of the earthworm Megascolex". J. Comp. Neurol. 94 (1): 57-103. doi:10.1002/cne.900940104. PMID 14814220.
- ^ "Ch. 20: Nervous system". The insects: structure and function. Cambridge University Press. 1998. ss. 533-568. ISBN 978-0-521-57890-5.
- ^ Chapman, p. 546
- ^ Ko (2016). "Origins of human intelligence: The chain of tool-making and brain evolution" (PDF). Anthropological Notebooks. 22 (1): 5-22. 17 Ağustos 2016 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 26 Haziran 2021.
- ^ "A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia". Nature. 431 (7012): 1055-61. 2004. doi:10.1038/nature02999. PMID 15514638.
- ^ Davidson. "As large as you need and as small as you can'--implications of the brain size of Homo floresiensis, (Iain Davidson)". Une-au.academia.edu. 25 Nisan 2012 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Ekim 2011.
- ^ "4.04 - Energetics, Life History, and Human Brain Evolution", Evolution of Nervous Systems (Second Edition) (İngilizce), Oxford: Academic Press, 1 Ocak 2017, ss. 51-62, ISBN 978-0-12-804096-6, 26 Haziran 2021 tarihinde kaynağından arşivlendi, erişim tarihi: 7 Nisan 2021 Birden fazla editör-name-list parameters kullanıldı (yardım); r
|ad1=eksik|soyadı1=(yardım)