Programlanmış hücre ölümü
Programlanmış hücre ölümü (PHÖ veya PCD), herhangi bir hücrenin, hücre içi bir programla ölmesinin planlaması ve gerçekleştirilmesidir.[1]
Nekrozdan farklı olarak, ölen hücrelerin sonucunda dokuda akut hasarlanma ve enflamasyon yanıt oluşturulmaz. Programlanmış hücre ölümü, genellikle canlının yaşamı boyunca yarar sağlayan düzenli bir süreçtir. PHÖ, bitkiler ve Metazoa gruplarının doku gelişimleri sırasında esasi fonksiyonları sergiler.
- Apoptozis ya da Tip I hücre-ölümü
- Otofajik ya da Tip II hücre ölümü (sitoplazmik - hücre çekirdeğinin yok edilmesinden önce özel bir sırayla büyük vakuollerin organelleri yemesiyle bilinen süreç.)[2]
Programlanmış hücre ölümünün bu iki şeklinden başka yollarda bulunmuştur.[3] apotoptik olmayan programlanmış hücre ölümü olarak bilinen (ya da "kaspas-bağımısz programlanmış hücre ölümü" veya "nekroz benzeri programlanmış hücre ölümü") bu alternatif ölüm yolları apoptozis gibi etkilidir, arka planda işlem görürler ve PHÖ'nün ana tiplerini oluştururlar.
Bitki hücrelerinde kısmen otofajik hücre ölümüne benzer şekilde PHÖ görülür. PHÖ'nün bazı temel özellikleri bitkiler ve Metazoada büyük oranda korunmuştur.
"Programlanmış hücre ölümü", 1964'te Lockshin ve Williams[4] tarafından böcek dokularının gelişimindeki ilişkilerde kullanılmıştır, ondan sekiz yıl kadar önce de apotozis keşfedilmiştir. O zamandan bu yana "PHÖ", bu iki terimden daha genel bir kullanım haline gelmiştir.
PHÖ, son zamanlarda gittkiçe artan ilgi ve araştırmaların konusu olmuştur. Bu eğilim, 2002'de Sydney Brenner (BK), H. Robert Horvitz (USA) ve John E. Sulston (BK)'ın Nobel Fizyoloji ve Tıp Ödülü'nü almasıyla da vurgulanmıştır.[5]
Bitki dokularında programlanmış hücre ölümü
Arabidopsis türlerinde tanımlanan "APL", düzenler vasküler dokuları düzenler[6] Bonke ve çalışma arkadaşları, vasküler bitkilerin uzun mesafeli taşıma yapan iki sisteminden biri olan ksilemde, çeşitli hücre tiplerinin "hücre duvarları kalınlıklarında ve programlanmış hücre ölümlerinde, döküntülerin dikkatlice bağlanmasının farklılıklar" içeridiğini belirtmişlerdir. Yazarlar, bitkilerdeki PHÖ ürünlerinin yapısal önemli bir rolü olduğunu da vurgulamıştır.
PHÖ'nün temel morfolojik ve biyokimyasal özellikleri, bitkiler ve hayvanlar aleminde korunmuştur.[7]
Bitki hücrelerin spesifik çeşitleri eşsiz hücre ölümü programlarını her nasılsa başardıklarını not düşmek gerekir. Hayvan apotozisiyle birlikte bunlar yaygın özelliklerdir -örneğin çekirdek DNA'sı degredasyonu- fakat bitkiler ksilemin trakeid elemanlarındaki vakuolün çökmesiyle tetiklenen hücre çekirdeği degredasyonu gibi kendi özelliklerine sahiplerdir.[8]
Oxford Üniversitesinden bitki bilimciler, Janneke Balk ve Christopher J. Leaver, ayçiçeği bitkisinin hücrelerindeki mitokondriyal genomdaki mutasyona ulaşmayı başarmışlardır. Bu araştırmanının sonucu olarak mitokondrinin vasküler bitkilerin ve diğer ökaryotik hücrelerin PHÖ'de temel rol oynadığı ileri sürülmüştür.[9]
Polenlerde kendini döllenmenin PHÖ ile önlenmesi
Polenizasyon sırasında bitkiler kendilerini döllemenin engellenmesinde önemli bir anlamı olan "kendiyle uyuşmazlığı" (SI) yürütürler. Gelincik bitkisi üzerinde yapılan araştırmalar polenlerin bulunduğu yer olan pistillerdeki proteinlerin polenle etkileştiğini ve örneğin kendi poleniyle uyuşmazlıkta PHÖ'nü tetiklediğini göstermiştir. Araştırmacılar Steven G. Thomas ve Veronica E. Franklin-Tong bir de, PHÖ'ce izlenen polen tüpü gelişiminin hızlı inhibisyonunun yanıt gerektirdiğini de bulmuşlardır.[10]
Mycetozoa'da programlanmış hücre ölümü
Sosyal tabaka küfü Dictyostelium discoideum eğer uniselüler formundaysa avcı amipe benzer bir davranış sergilerken, gelecek nesli verecek olan sporlarını dağıtırken hareketli bir sümüklüböceğe benzer bir özellik sergiler.[11]
Bitkinin ölü hücrelerden oluşan sapı, bir çeşit PHÖ olan otofajik hücre ölümünün bazı özelliklerini paylaşır: hücrelerin içide oluşan büyük vakuoller, kromatin yoğunlaşmasının bir aşaması fakat DNA'nın hiç parçalanmaması.[12] Ölü hücrelerce bırakılan kalıntıların yapısal rolü bitki dokusundaki PHÖ'nin ürünlerini anımsatır.
D. discoideum, günümüzden milyonlarca yıl önce ökaryotik atalarından meydana gelen olası bir dalın parçası olan sıvaşık bir küftür. Görünüşe göre, onlar yeşil bitkilerin, mantarların ve hayvanların atalarından sonra farklılaşmışlardır. Fakat evrim ağacındaki yerlerine göre gerçek şudur; PHÖ basitçe D. discoideumun 6. kromozomunda saklanıp, ona ek bir anlam katar: apoptozisin özelliklerinden olan kaspaslara bağımlı olmayan gelişimsel PHÖ yolunun çalışmasına izin verir.[13]
PHÖ'nün evrimsel kökeni
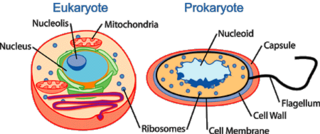
Biyologlar uzun zaman mitokondrinin daha büyük ökaryotik hücrelerden endobiyontları da kapsayan bakterilerden kökenlendiğinden şüphelendiler. Lynn Margulis 1967'de bu teorinin kazananı oldu, gerçek dünyaca kabul edildi.[14] Bu teorinin inandırıcı olayı, mitokondrinin kendi DNA'sına sahip olması ve genler ve replikasyon aygıtlarıyla donatılması gerçeğinde yatar. Bu evrimsel adım, enerji üreten bakterileri yutmaya başlayan ilkel ökaryotikler için daha riskli ve zıt olarak proto-ökaryotik konaklarına saldırmalarıyla başlayan mitokondrilerin ataları için çok tehlikeli olacaktır. Bu, insan beyaz kan hücreleri ve bakteriler arasındaki süreç günümüzde hala devam eder. Zamanının çoğunda, saldırgan bakteriler beyaz kan hücrelerince yokedilir, bununla beraber; prokaryotların hasarlanması sonucu enfeksiyon diye bilinen sonuçla bilinen kimyasal savaş için çok da olağanüstü değildir. Bu nadir evrimsel olaylardan birini günümüzden yaklaşık 2 milyon yıl önce kesin ökaryotlar ve sadece varolmayıp, enerji üreten fakat simbiyozimden karşılıklı yarar sağlayan prokaryotlar için mümkün yapar.[15] Mitokondriye sahip ökaryotik hücreler ölümle yaşam arasında bir çizgide canlı tutulur, çünkü mitokondri kendi hücre intiharını tetikleyebilecek molekülleri hala içermektedir.[16] Bu süreç günümüzde sadece programlandığı zaman yavaş olarak gelişir. Hücrelere verilen kesin sinyallerle (komşulardan uyarı, stres ya da DNA hasarı gibi) mitokondri biyokimyasal kaskada neden olup, hücre ölümünü tetikleyen kaspas aktivatörlerini salar. Bu şekilde, hücre intiharı mekanizması günümüzde yaşamlarımızda var olan önemli bir ayrıntıdır.
Canlılarda PHÖ
Kaynakça
- ^ Engelberg-Kulka H, Amitai S, Kolodkin-Gal I, Hazan R (2006). "Bacterial Programmed Cell Death and Multicellular Behavior in Bacteria". PLoS Genetics. 2 (10). ss. 1518-26. 11 Temmuz 2012 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Ekim 2020.
- ^ Lawrence M. Schwartz; ve diğerleri. (1993). "Do All Programmed Cell Deaths Occur Via Apoptosis?". PNAS. 90 (3). s. 980. 8 Kasım 2007 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Şubat 2008.;and, for a more recent view, see W. Bursch; ve diğerleri. (2000). "Programmed Cell Death (PCD): Apoptosis, Autophagic PCD, or Others?". Annals of the New York Academy of Sciences. Cilt 926. s. 1.
- ^ Kroemer G, Martin SJ (2005). "Caspase-independent cell death". Nat Med. 11 (7). ss. 725-30.
- ^ Richard A. Lockshin, Carroll M. Williams (1964). "Programmed cell death—II. Endocrine potentiation of the breakdown of the intersegmental muscles of silkmoths". Journal of Insect Physiology. 10 (4). ss. 643-649. 28 Şubat 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Şubat 2008.
- ^ "2002 Nobel Prize in Medicine or Physiology". 29 Ekim 2002 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Şubat 2008.
- ^ Martin Bonke et al., published in Nature Vol. 425, Nov. 13, 2003, p. 181.
- ^ Mazal Solomon, et al.: "The Involvement of Cysteine Proteases and Protease Inhibitor Genes in the Regulation of Programmed Cell Death in Plants", The Plant Cell, Vol. 11, 431-444, March 1999. See also related articles in The Plant Cell Online, [1] 16 Kasım 2007 tarihinde Wayback Machine sitesinde arşivlendi.
- ^ See Jun Ito and Hiroo Fukuda: "ZEN1 Is a Key Enzyme in the Degradation of Nuclear DNA during Programmed Cell Death of Tracheary Elements", The Plant Cell, Vol. 14, 3201-3211, December 2002.
- ^ "The PET1-CMS Mitochondrial Mutation in Sunflower Is Associated with Premature Programmed Cell Death and Cytochrome c Release", The Plant Cell, Vol. 13, 1803-1818, August 2001
- ^ Thomas, and Franklin-Tong: "Self-incompatibility triggers programmed cell death in Papaver pollen", Nature Vol. 429, 20 May 2004, p. 305.
- ^ Crespi B, Springer S: "Ecology. Social slime molds meet their match" Science 2003 Jan 3;299(5603):105-6 [2] 17 Nisan 2022 tarihinde Wayback Machine sitesinde arşivlendi..
- ^ See the article by Levraud et al.: "Dictyostelium cell death : early emergence and demise of highly polarized paddle cells", The Journal of Cell Biology Vol. 160, 7, p. 1105 [3] 8 Kasım 2007 tarihinde Wayback Machine sitesinde arşivlendi..
- ^ See also Roisin-Bouffay et al.: "Developmental Cell Death in Dictyostelium Does Not Require Paracaspase", The Journal of Biological Chemistry Vol. 279, 12, p. 11489, 19 March 2004 [4] 8 Kasım 2007 tarihinde Wayback Machine sitesinde arşivlendi..
- ^ see "The Birth of Complex Cells", by Christian de Duve, Scientific American Vol. 274, 4, April, 1996
- ^ See "Ancient Invasions: From Endosymbionts to Organelle", by Sabrina D. Dyall et al., Science Vol. 304 p. 253, 9 April. 2004 [5] 17 Nisan 2022 tarihinde Wayback Machine sitesinde arşivlendi..
- ^ see Chiarugi and Moskowitz, in Science 297, p. 200 [6] 17 Nisan 2022 tarihinde Wayback Machine sitesinde arşivlendi.
Dış bağlantılar
- www.caspases.org25 Ağustos 2014 tarihinde Wayback Machine sitesinde arşivlendi.
- www.celldeath-apoptosis.org 21 Şubat 2021 tarihinde Wayback Machine sitesinde arşivlendi.