Dinozor
| Dinozor | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Çeşitli dinozor iskeletleri. Soldan saat yönüne doğru: Microraptor gui (kanatlı bir theropod), Apatosaurus louisae (dev bir sauropod), Edmontosaurus regalis (bir ördek gagalı ornithopod), Triceratops horridus (boynuzlu bir ceratopsian), Stegosaurus stenops (zırhlı bir stegosaur), Pinacosaurus grangeri (zırhlı bir ankylosaur) | |||||||||
| Biyolojik sınıflandırma | |||||||||
| |||||||||
| Alt gruplar | |||||||||
| |||||||||
Dinozor[a] (Dinosauria), ilk olarak Mezozoyik zamanda ortaya çıkan ve yaşayan tek üyeleri kuşlar olan arkozor sürüngen grubu. Dinozor adı, Richard Owen tarafından Grekçe "korkunç" anlamına gelen deinos ve "kertenkele" anlamına gelen sauros sözcüklerinin birleştirilmesinden oluşur.[7] Dinozorlar, 243 ile 233 milyon yıl önce Geç Triyas döneminde ortaya çıkmış ve 66 milyon yıl önce kuşlar dışındaki tüm türlerinin soyu tükenmiştir. Dinozorlar, sıcakkanlı ve soğukkanlı arası özellikler gösteren mezoterm canlılardı. Mezozoyik'te oldukça başarılı biçimde tüm kıtalara yayılan dinozorlar çok farklı nişleri doldurdu. Yapılan araştırmalarda şimdiye kadar yaklaşık olarak 1.000 kadar kuş olmayan dinozor türünün yaşadığı belgelendi. Bugün dinozorların evrimsel olarak devamı olan kuşların 11.000 kadar türü vardır ki bu, yaşayan memelilerin tür sayısının (~6000) yaklaşık iki katıdır. Dinozorların nasıl yaşadığı, ne kadar çeşitlendiği ve ekosistemdeki yerleri kadar kuş olmayan dinozorların nasıl yok olduğu da bilim camiasını uzun zamandır meşgul etmektedir. Kuş olmayan dinozorları yeryüzünden silen yok oluşun aşamalı mı, yoksa yerbilimsel olarak katastrofik (ani) mi olduğu tartışmalıdır.[8]
Dinozorlarla ilgili bir diğer konu da onların büyük çeşitlilik gösteren boyutlarıdır. Bu canlılar, yerküreyi titreten 65-75 tonluk[9] Argentinosaurus gibi dev titanozor sauropodlardan en küçük yaşayan dinozor olan Arı sinek kuşuna değin boyut konusunda çok geniş bir yelpaze sunar. Farklı dinozor grupları farklı metabolizma hızları ve büyüme stratejileri geliştirdi, sauropodlar devleşip ısı kaybını aza indirirken, daha küçük bedene sahip gruplar da ısı kaybını önlemek için tüy barındırmış olabilir. Dinozorların ayırt edici fiziksel özelliklerinden birisi beden duruşlarıdır, yapılan çalışmalar dinozorların atalarının bipedal (iki ayak üstünde) olduğunu ileri sürdü. Yaşayan kertenkele türlerindeki bipedal duruşun evrimsel olarak koşucu özellikleri artırıcı bir adaptasyon olduğu biliniyor, bundan yola çıkarak kimi dinozorlarda da gerek avlanma gerekse avcılardan kaçmak için bipedal hareket biçiminin evrimleştiği düşünüldü.[10]
Dinozorların evrimsel tarihi temelinde yerküre tarihidir, özellikle Dünya'nın geçirdiği büyük değişiklikler farklı dinozor ailelerinin veya aynı aileler içinde farklı cinslerin evrimleşmesine yol açtı. Bunun en önemli örneklerinden biri Kretase dönemi Kuzey Amerika'sında karaları ayıran Batı İç Denizyolu'dur ve yaşanan coğrafi izolasyon yoluyla Laramidya ve Appalachia karalarında ayrı cinsler oluştu.[11]
Keşif

Dinozorların ilk olarak ne zaman toplumların ilgisini çektiği bilinmiyor ancak Antik Çağ'dan başlayarak belli varsayımlarda bulunulabilir. Bazı tarihçiler ve paleontologlar Geç Kretase döneminin Kampaniyen aşamasında yaşamış Protoceratops'un fosilleri bulunduğunda, bunların Greko-Romen kültürde yansımasını bulmuş olabileceğine inanıyorlar. Bunun yanı sıra Antik Çin kültüründeki ejderha motifinin de dönemin insanlarının bulduğu dinozor kemiklerinden kaynaklandığı ileri sürüldü.[12] Muhtemelen bu kalıntıların bulunması ve onların dönemin insanlarınca masalsı yaratıklar olarak adlandırılması söz konusu olabilir (Griffon, anka kuşu vb).[13] Bununla birlikte 2024 yılında Witton ve Hing'ın yaptığı çalışma, önceden ileri sürülen Protoceratops'un kalıntılarının altın arayan Asyalı göçebelerce bulunduğu ve daha sonra griffon adlı mitolojik varlığın yaratılmasını esinlediği ve bunun ticaret ile antik dünyada yayıldığı varsayımına karşı bir görüş ileri sürdü. Çalışmanın ana argümanları fosil kalıntılarının yakınında altın yataklarının bulunmaması ve griffon sanatının coğrafi yayılımının Orta Asya'dan kaynaklanma ve sonradan Batı'ya yayılma iddiasına uymamasıydı.[14]
Yeni Çağ dolaylarında artık modern bilimin ayağa kalkmaya başladığı zamanlarda eksik keşifler yapılmıştır. 1676'da Oxford Üniversitesi'nde görevli bir rahip olan Robert Plot, bir dinozora ait olduğu bugün bilinen bir uyluk kemiğini Roma döneminden kalma savaş filine ait bir kalıntı sanmıştır. Örnekler çoğaltılabilir; ancak dinozorların ve doğa bilimlerinin doruk noktası 19. asrın başlarıdır. İlk olarak dinozorların ciddi bilimsel araştırmaların konusu olması İngiliz doğa bilimci Richard Owen'in katkısıyla oldu, çünkü dinozor terimini ortaya koyan kendisiydi.[15] Bilimsel anlamda incelenen ilk dinozor fosili ise Megalosaurus'tur, bu orta Jura dönemi etçili, William Buckland tarafından bulunmuş ve tanımlanmıştır.[16] Tanımlanan ikinci dinozorsa 1822 yılında Gideon Mantell'in keşfettiği İguanodon'dur, cins 1825 yılında Mantell tarafından resmi olarak tanımlanmıştır.[17] Bilimsel anlamda son 200 yılda bir avuç eksik numuneyle başlayan dinozorları tanımlama süreci bugün en az 800 cins ve neredeyse 1000 türle çok büyük bir sayıda temsil edilmeye başladı. Bu geniş canlılar grubunun da hâlâ daha yeni bulgularla değişebilecek bir evrim süreci vardır.[18]
Avrupa'da bu gelişmeler ve keşifler yapıldıktan sonra paleontoloji bilimi Amerika kıtasındaki çalışmalarla daha da genişlemiştir. Bilhassa Avrupa'da eğitim almış, ama saha çalışmalarını Kuzey Amerika'da yapan iki önemli bilimsel kişilik Othniel Charles Marsh ve Edward Drinker Cope sahneye çıkmıştır. Bu önemli keşifler git gide ekol hâline gelmiş ve ikili etrafında rekabete dayalı bir bilimsel hareket başlamıştır. Hem Cope hem de Marsh arazi gezilerine çıkarak fosil ararlarken; bir süre sonra arazi çalışmaları için işçi ekipleri kurup başlarına asistanlarını geçirmişlerdir. İlerleyen zamanlarda, bulunan ve ulaştırılan fosilleri ofislerinde inceleme ve tasnif etme işlerini yürütmüşlerdir. Bu ikilinin rekabeti ise fikir ayrılıkları ve fosil alanları ile ilgili çatışmalar nedeniyle kavgaya dönüşmüş olup 1890 yılının ocak ayında bu kavga gazete manşetlerine düşmüş ve kamuoyunun taraf olduğu bir hadiseye dönüşmüştür. Aralarındaki kavga, düşmanlık boyutuna vardı ve rüşvet, hırsızlık ve fosillere zarar vermeye kadar uzanan kötü eylemlere sahne oldu.[19] Bu bilimsel tartışma ise bilim tarihine "Kemik Savaşları" (İngilizce: Bone Wars) olarak geçmiştir.[20]
Paleobiyoloji

Boyut
Dinozorlar pek çok özellikleri noktasında yüksek çeşitlilik yakaladılar. Boyut bu özelliklerin başında gelmektedir. Memeliler ile karşılaştırmak gerekirse ortalama memeli boyutu köpek ölçülerindeyken; ortalama dinozor boyutu ayı ölçülerine karşılık gelir. Tüm dinozorlar, boyut konusunda dikkate değer bir evrimsel öykü sunar, Mezozoyik dönemde dinozor beden kütlesinin modu 1-10 ton arasındaydı.[21] Günümüzde tamamıyla karada yaşayan en büyük sürüngenler dev kaplumbağalar ve monitör kertenkelelerdir ve 1 tona yaklaşamazlar, ancak büyük etçil dinozorlar 6-10 tona kadar ulaşabilen örneklere sahiplerdi. Geç Jura'nın Avrupa'sından bilinen Compsognathus 1-1,25 m uzunluğundayken,[22] Tyrannosaurus rex 12 metre uzunluğunda ve 8[23] ton kadar gelmekteydi. Sauropodlar için boyut konuşulacaksa devleşme söz konusu olmalıdır, örnek olarak 75 tona kadar ulaşabilen Argentinosaurus[9] ve 35 tonluk Supersaurus verilebilir. Bilinen en büyük ornithischian ve sauropod olmayan en büyük dinozor olan Shantungosaurus için de Gregory S. Paul, 15 m uzunluk ve 13 ton ağırlık tahmin etti.[24]
Ayrıca 2024 yılında yapılan bir çalışmada Tyrannosaurus cinsine yeni bir tür eklenmiştir. T. mcraeensis, T. rex'ten 6-7 milyon yıl önce Laramidya'nın daha güneyinde yaşadı ve kendisinden daha genç akrabasıyla yarışır boyutlara sahipti. T. mcraeensis'in 12 metre uzunluğa sahip olduğu düşünülüyor. Bu yeni türün keşfiyle Tyrannosauridlerin büyük boyutlara nasıl ulaştıkları ve kökenlerine ilişkin evrimsel tarihleri daha iyi anlaşılabilir.[25]
Dinozorlar aynı zamanda farklı beden yapılarının boyutu konusunda da uç örnekler sergilediler. İlginç örneklerden biri Mamenchisaurus'tur. Andrew Moore ve ekibi tarafından 2023'te yapılan çalışmaya göre 15 m uzunluğundaki boynuyla Mamenchisaurus'un şimdiye kadar bilinen en uzun boyna sahip canlı olduğu bulundu.[26]
Bergmann kuralı zoolojide hayvanların boyutlarıyla yaşadıkları enlem arasındaki ilişkiyi konu eden bir ekolojik kuraldır. Wilson ve ark.'nın (2024) yaptıkları çalışmada bu kural Mezozoyik'te yaşamış dinozorlar ve memeliler ile onların torunları olan günümüz kuşları ve memelilerine uygulandı. Özellikle son yıllarda keşfedilen Kuzey Kutup Dairesi çevresinde yaşamış Nanuqsaurus gibi dinozorların da ele alındığı çalışmada beden büyüklüğünün yaşanılan yerin enlemiyle bir ilgisiyle olmadığı bulundu. Kuzey Amerika'nın daha alt enlemlerinde yaşamış dinozorlarla onların Arktik çevrelere yakın yaşamış akrabalarının beden büyüklüklerinin benzer olduğu görüldü.[27]

Metabolizma
Omurgalılar dünyasında temel olarak iki tip enerji üretim sistemi metabolizmayı ve canlı yaşamını yönetir. Bunlardan biri yüksek metabolizma hızına sahip memeli ve kuşlardaki enerji sistemiyken; diğeri düşük metabolizma hızına sahip sürüngen, amfibi ve balıklarda görülen enerji üretim sistemidir. Dinozorların genellikle dik bacaklara[22] sahip olması onların da memeli ve kuşlar gibi etkin canlılar olduğu fikrini desteklemektedir. Bu noktada pelvisin (kalça kemiği) boyutu önemli bir belirteçtir, sürüngenlerde yavaş harekete olanak sağlayan ufak bacak kasları (uyluk kasları) yine görece küçük bir pelvise bağlanır ve bu durum kuş ve memelilerde tam ters durumdadır onların büyük pelvisleri vardır. Özellikle prosauropodlar, erken teropodlar gibi bazı ilkin dinozor grupları, küçük pelvislere sahiptir ve kimi çalışmalarda onların da düşük metabolizma hızına sahip olduğu düşünülmüştür, ancak özellikle avian dinozorlarda, gelişmiş teropod ve sauropodlarda, modern kuşlardaki gibi hava keselerinin bulunması, bazı kuş kalçalı dinozorlarda diyafram benzeri bir yapının korunması da dinozorların yüksek metabolizma hızına veya en azından sürüngenler ile kuş-memeli metabolizması arasında bir metabolizmaya sahip olduğunu gösterir. Ayrıca fosilleşmiş kemik kalıntıları da canlının yaşadığı süreçteki ısı değişimini gösterecek izotoplar barındırır, yapılan incelemelere göre dinozorlar genelde aynı ortamda yaşadıkları timsahgillerden daha duraylı (sabit) vücut sıcaklıklarına sahip olmuştur.[24]
Dinozorların vücut sıcaklıklarını dengelemek için sahip oldukları çeşitli adaptasyonları vardır. Büyük sauropod Camarasaurus ağır yapısı nedeniyle kolayca ısı kaybedemez ve aşırı ısınan kafatasını burun boşluklarında ve ağzında yer alan ısı transfer alanlarındaki kan damarlarıyla fazla ısıyı atarak soğutur. Yine Euplocephalus'ta döngüsel burun boşlukları vardır ve geniş kan damarları burada fazla ısıyı uzaklaştırır. Orta boy bir teropod olan Majungasaurus çenesiyle bağlantılı bir sinüs sistemine sahiptir, hayvan ağzını açtığında bu sinüslere hava dolar, sonra sinüsleri çevreleyen kan damarları havayı ısıtır ve fazla ısısını havaya kaybeden serin damarlar beyne gitmektedir. Hayvan ağzını kapatınca da ısınan hava burun ve ağızdan dışarı atılır.[28]
Son zamanlardaki çalışmalarda Tyrannosaurus rex'in kafatasındaki deliklerin farklı işlevleri olabileceği bulundu. İlk olarak hayvanın kafasının soğumasına yardım ettiği düşünüldü, diğer olasılıklar da kafatasının ağırlığını azaltan veya daha iyi bir işitme duyusuna sahip olmasına yardım eden bir adaptasyon sayılmasıdır.[29]
Dinozorların büyüme hızları da metabolizma incelemelerine konu edilir. Sauropodların evrimsel olarak boyutlarını büyütme eğilimi günümüzdeki büyük memelilerle karşılaştırılmıştır. Örnek olarak, beş yaşında bir afrika fili 1 ton kadar gelirken; aynı yaştaki Apatosaurus'un ise 20 tona ulaştığı biliniyor. Bu büyüme hızı yüksek görünse de yaşayan balinalarla karşılaştırılınca çok yüksek olmadığı görülmektedir. Yine 30 tonluk gri balina, 20,700 kg/gün büyüme hızı yakalarken Apatosaurus 14,460 kg/gün büyüme hızına sahiptir. Yaşamış en büyük kara hayvanı olduğu tahmin edilen Argentinosaurus'un da 55,638 kg/gün'lük büyüme hızıyla, 66,00 kg/gün'lük büyüme hızına sahip gelmiş geçmiş en büyük hayvan mavi balinanın gerisinde kalır.[30] 2023 yılında D'emic ve meslektaşlarının yaptığı çalışmada farklı teropod dinozorların büyüme hızları ve süreleri belirlendi. Uyluk ve kaval kemiklerindeki büyüme halkaları üzerinde yapılan incelemeler sonucu Spinosaurus, Tyrannotitan, Acrocanthosaurus, Majungasaurus gibi cinslerin Tyrannosaurus'tan daha yavaş büyüdüğü saptandı. Mapusaurus'un da yılda % 200 oranında kütlesini artırarak hızlı büyüdüğü belgelendi.[31]
2024 yılında Rogers ve arkadaşları tarafından yapılan bir çalışmaya göre Triyas döneminde yaşayan ilk dinozorların ve onlarla yan yana yaşayan diğer sürüngenlerin benzer olarak hızlı ve sürekli büyüdüğü ortaya kondu. Ischigualasto Formasyonunda yaşayan bir dizi dinozor ve sürüngenin uyluk kemiklerinin histolojisi üzerine yapılan araştırma ile erken dinozorların Jura ve Kretase dönemlerindeki torunlarından daha sürekli büyüdüğü de keşfedildi.[32]
Üreme ve davranış
Dinozorlar da tıpkı kuşlar ve sürüngenler gibi yumurtlayarak üremiştir ve bu durum fosilleşmiş yumurta ve hatta embriyo kalıntılarıyla belgelenmiştir. Troodon, Maiasaura, Oviraptor ve Hypacrosaurus cinslerine ait yumurtalar bulunmuştur ve en tam yumurta grubu da Troodon'a ait olanlardır. Yuva yapımı ve dinozorların kuluçkaya yatmasıyla ilgili ciddi kalıntılar ortaya çıkmıştır, en ünlü örnek yuvası üstüne kapanan Oviraptor fosilidir. Oviraptor kalıntıları ilk keşfedildiğinde bir Protoceratops yuvasına olan yakınlığı dolayısıyla yuvadan yumurtaları çaldığı düşünülerek bu canlıya yumurta hırsızı anlamına gelen Oviraptor adı verilse de 1993'de Mark Norell, ilgili yumurtaların birinde bulduğu embriyo inceleyerek yumurtaların Oviraptor'a ait olduğunu ortaya çıkardı ve bu dinozorun hırsızdan çok bir ebeveyn olduğu anlaşıldı.[33] Ayrıca Troodon yumurtalarının bulunduğu yuvanın kenarlarında topraktan 12 cm'lik bir bariyer bulunmuştur.[34]
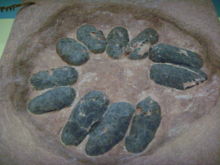
Yavru bakımı konusunda kuş kalçalı dinozorlardan gelen fosil kanıtları bulunmuştur, özellikle Maiasaura adlı ördek gagalı dinozorun yavrularına tıpkı modern kuşlarda görüldüğü gibi baktığı anlaşılmıştır.[34][35]
Dinozorların bazı türlerinin sürüler halinde yaşamış olabileceği ileri sürülmektedir, özellikle toplu fosil alanları bu konuda en önemli kanıtları oluşturmaktadır. Hadrosauridler ve ceratopsidler yuva alanlarında veya normal yaşamlarında toplulukla hareket ediyorlardı.[36] Hem genç hem de yetişkin dinozorların fosillerinin bir arada bulunması veya toplu dinozor mezarlarının ortaya çıkmasıyla dinozorların toplu yaşamları konusunda daha net açıklamalar getirilebilmiştir.[35] Wyoming'de bulunan beş Triceratops bireyine ait 1200 kemik ve kemik parçası üzerinden yapılan bir çalışmaya göre de bu dinozorların sürü davranışı gösterip göstermediği tartışıldı ve makalesinin de yazımına başlandı. Yüzlerce Triceratops dişinin incelenmesine dayanrak da bu beş dinozor için ortak olan toplu göçebe bir yaşam biçimi ileri sürüldü.[37]
Dinozorların özel vücut yapıları da türler arası ve tür içi iletişimde önemli yer tutmaktadır. Özellikle boynuzlar, dikenler veya kemiksi yapılar bu oluşumlara örnektir. Boynuzların eşleri cezbetmek veya avcılardan korunma amaçlı kullanıldığı düşünülür, tıpkı stegosauridlerdeki dikenlerin kullanıldığı gibi. Ayrıca yavru-yetişkin iletişiminde de fizyolojik farklar rol oynamaktadır, örnek olarak yavru hadrosauridlerin yetişkinlerden daha farklı bir iskelet yapısı vardır.[35] Teropod dinozorlarda da bireylerin kendi aralarında belli etkileşimlere girdiği bazı fosil kanıtlar üzerinden anlaşıldı. Özellikle fosil teropod çeneleri ve kranyumlarındaki yara, delik veya sıyrık izleri bu canlıların birbirleriyle av, yaşam alanı, yuva yeri veya çiftleşme hakkı için kavga ettiğini tezini doğruladı.[38] Teropodlar arasında kurak günlerde yamyamlık görüldüğü hipotezi de birtakım fosil materyallerde bulunan travma izleri üzerinden açıklandı. Mygatt-Moore Quarry'deki kayalar Geç Jura yaşlı Morrison formasyonundan gelen dinozorların fosil topluluğunu içerir. Buradan gelen teropod fosillerinde başka bir teropod tarafından bırakılan izlere rastlandı. Diş izlerini Allosaurus, Ceratosaurus veya daha büyük olan Saurophaganax gibi cinslerden biri bırakmış olabilir.[39]
Dinozorlar çağından kalan av avcı ilişkisinin somut bir kanıtı 1971'de Moğolistan'ın Geç Kretase yaşlı katmanlarında bulunan ve Savaşan Dinozorlar olarak ünlenen fosil kalıntılardır. Bu fosil örnek, bir Protoceratops ve Velociraptor arasındaki mücadeleyi betimlemektedir. Daha da ötesi bu iki canlının olası bir kum fırtınası veya kum tepesi çökmesi sonucu canlı canlı tam dövüşürlerken gömülmesidir.[40] Dinozorların dünyasındaki davranış kalıplarına ilişkin çarpıcı bir örnek sunar.[41]

Ünlü spinosaurlardan biri olan Irritator ile ilgili yapılan bir çalışmada dinozorun kafatası ve alt çenesinin duruşu ve yapısı konusunda yeni keşifler yapıldı. Yarım daire kanalları ve oksipital kondilin yönelimlerine dayanarak hayvanın başının yaklaşık 45° kadar öne eğik durduğu iddia edildi. Bunun yanı sıra, Irritator'un alt çenesindeki baskıcı kasların, paroksipital çıkıntının arka-yan yüzeyi ile retroartiküler çıkıntının posterodorsal yüzeyi arasında uzanarak bu canlının alt çenesinin güçlü ve hızlı bir açılma hareketine izin verdiği bulundu. Günümüzdeki pelikanlar gibi alt çenelerini beslenme sırasında genişlettikleri ileri sürüldü.[42][43]
Yakın zamanda Çin'in Guizhou eyaletindeki Ziliujing Formasyonunda 3 yetişkin dinozor ve yumurta kümeleri bulundu. Yapılan çalışmada bu yumurtaların kabuğunun dokusunun derimsi olduğu anlaşıldı ve bu özelliğin arkozor atalarından gelen bir özellik olabileceği ileri sürüldü. Yumurta kabuğu kalınlığındaki artışın da teropod ve sauropodomorf evriminde erkenden gerçekleştiği bulundu.[44]
Paleopatoloji
Fosil materyallerin araştırılmasıyla günümüz hayvanlarını etkileyen hastalıkların soyu tükenmiş canlılarda da olduğu ortaya kondu. Alberta, Kanada'da 77 milyon yıllık bir fosil yatağında bir Centrosaurus sürüsüne ait fosil kalıntılar bulunmuştur. Bu dinozorların toplu halde fosilleşmesi ani gelişen fırtınanın yarattığı selin hepsini boğmasıdır, ama bu fosil yatağındaki bir Centrosaurus'un kemikleri çok önemli bir bulguya işaret ediyordu. Ekhtiari ve ark. yaptığı çalışmada bir Centrosaurus bireyinin kaval kemiğinde osteosarkoma adlı kötü huylu bir kemik tümörünün bulunduğu ortaya konmuştur. Bu hasta dinozor erkek ve 19 yaşında bir bireydi.[45]
Başka bir çalışmada Montana Dinozor Müzesinden Cary Woodruff, 145 milyon yıllık Diplodocid benzeri bir sauropodun boyun kemiklerinde garip, brokoli benzeri büyümeler gözlemledi. Yapılan BT taramaları ve X-ray incelemeleri ile bu oluşumların solunum yolları enfeksiyonu sonucu geliştiği ortaya konmuştur. Uzmanlar ilgili dinozorun, olasılıkla ateş, akıntı ve öksürükten muzdarip olduğunu düşünüyorlar. Bu keşif, kuşlara özgü solunum yolu enfeksiyonlarının kuş olmayan dinozorlarda görüldüğü ilk vaka olma özelliğini taşıyor.[46] "Dolly" adı verilen dinozorun günümüzde de var olan hastalıklardan birisini göstermesi bu tip hastalıkların evrimsel tarihine ışık tutacaktır.[47]

Baiano ve ark.'nın (2024) yaptığı bir çalışmaya göre Tetanurae olmayan teropodlarda ilk kez patolojik bir durum keşfedildi. Abelisaurid olan Aucasaurus'un kuyruk omurları ile hemal yayının sıkı şekilde birbirine bitişik olduğu bulundu. Bilgisayarlı tomografi ve morfolojik incelemeler kuyruk omurlarının büyümelerini tamamlamadığı ve birbirlerinden ayrılmadığını ve böylece bunun doğuştan bir şekil bozukluğuna işaret ettiğini gösterdi. Yine aynı çalışmada bir başka abelisaurid Elemgasem'in kuyruk omurlarında patolojik koşullara rastlandı. Omur gövdesinin eklem yüzeyinin sağ kenarında kemik büyümesi saptandı ve omur gövdesi arasındaki belirgin intercentrum boşlukları bu hayvanın artritten muzdarip olduğunu ortaya koydu.[48]
Sınıflandırma
Dinozorlar hakkındaki ilk bilimsel açıklama çabaları Richard Owen'dan gelmiştir ki, bu canlılara genel dinozor adını veren de oydu. Richard Owen başlangıçta Iguanodon, Megalosaurus ve Hylaeosaurus olarak adlandırdığı 3 cins üzerinden bir sınıflandırma sistemi kurmaya çalıştı, hatta 1851 yılındaki ünlü sergi için Crystal Palace Parkı'nda Owen'ın isteğiyle heykeltıraş Waterhouse Hawkins ölçekli olarak heykeller yaptı, o dönem dinozorların dev kertenkeleler olduğu düşüncesi hâkim olduğu için sergilenmek üzere, yaşayan sürüngen türlerinin heykelleri de yapılmıştı. Dönemin bilim dünyası, yeni fosillerin ışığında Mezozoik sürüngen çeşitliliğinin günümüzden çok olduğu kanaatine varmıştı. Yeni sınıflandırma önerileri peşi sıra geldi. Önce T. H. Huxley 1868'de yeni bir takım önerdi. Ona göre Ornithoscelida adlı bir takım tasarlanmalıydı, zira bu canlıların kuşlara benzeyen çok yönü vardı. Bu takımı ikiye ayırdı ve ilkini dinosauria yapıp Iguanodontia ve Megalosauria grubunu ve Scelidosaurus cinsini bu alt takıma koydu ve kuş benzeri küçük etçil Compsognathus da diğer alt takıma alındı.[18]
Bir süre sonra tabii ki yeni fikirler üretildi; ancak en yetkini 1887'de İngiliz biyolog H.G. Seeley'nin[49] hipoteziydi. Seeley, fosili bulunan dinozorların kemik yapılarını incelediğinde ayırıcı iki tip kalça kemiği şekli görmüştür. Bunlardan bir kısmı kuş kalçasına benzer bir yapıdayken; diğerleri kertenkele kalçasına benzer yapıdadır. Ayrımı takım seviyesinde terimleştirmiştir: (Ornithischia) kuş kalçalılar ve (Saurischia) kertenkele kalçalılar. Bu takımları da alt takımlara bölmüştür. Ornithischia dört alttakım içeriyordu: Ornithopoda (Iguanodon ve benzerleri), Stegosauria (plakalı türler), Ankylosauria (Hylaeosaurus ve diğer zırhlı türler) ve Ceratopsia (Kuzey Amerika'da yeni keşfedilen boynuzlu türler). Ayrıca kuş kalçalı dinozorların kalça özellikleri ve beslenme gibi ortak biyolojik karakterleri Geç Kretase zamanına dek korunmuştur.[50]
Seeley'nin ikinci takımı Saurischia ise tüm teropod (etçiller) cinslerini içerdiği gibi büyük otçulları (Cetiosaurus, Brontosaurus) da içeriyordu.[18] Bu devrimsel sınıflama dinosauria grubu üzerinde birtakım şüpheler doğurdu zira iki takıma bölünen bu canlıların ortak şemsiyesi dinosauria grubu olabilir miydi, yoksa tamamen farkı canlılar mıydı. Bu belirsizlik bir asır daha sürse de artık Dinosauria'nın ortak özellikler paylaşan bu canlıların doğal bir şemsiyesi olduğunu biliyoruz. Bundan bir sene sonra Belçika'nın Benissart kasabasında bir kömür madeninin işletmesi yapılırken; kazara bir düzine eklemli ve tam Iguanodon fosili bulundu. Bu muazzam bir keşifti ve Belçika Kraliyet Doğa Bilimleri Enstitüsü uzun bir süre bu çalışmaları sürdürdü ve 1883'te bu Iguanodon sürüsü sergilendi. Bu hayvanların sürü hâlinde dolaştığı ve bazılarının bipedal (iki ayak üstünde) hareket ettiği anlaşılabildi. Dinozorların sınıflandırılması konusunda önemli bir nokta da dinozorların kökeni meselesidir. Dinozorların ataları için en temel açıklama erken Triyas döneminde yaşayan arkozor cinsini işaret eder: Euparkeria. Geç Triyas'ın erken dönemine (237-201 milyon yıl önce) tarihlenen bazı arkozor cinsleri de dinozorların atası olarak kabul edilmektedir: Lagosuchus, Lagerpeton. Hatta Herrerasaurus ve Eoraptor gibi, bazı bilim insanlarının dinozor kabul ettiği, kimilerininse dinozor benzeri saydığı cinsler bulunmaktadır.[50]




Paleoekoloji
Dinozorlar ve bitkiler
Dinozor araştırmalarında bitki-dinozor etkileşimini gösteren doğrudan fosil kanıtlar (korunmuş dinozor dişlerindeki bitki izleri gibi) olmamasına[51] karşın bitkisel dünyaya ilişkin incelemeler ve hipotezler yapmak zorunludur, çünkü bitkiler Mezozoik Çağ'da çoktan kara yaşamına egemen olmuş ve büyük bir çeşitlilik arz etmeye başlamışlardı. Bitkilerin iki anlamda dinozorlar üzerinde büyük etkisi olmuştur. Bunlardan ilki besin olarak onların ekosisteminde temel üretici rolü üstlenmişler, ikinci olarak da evrimsel olarak onların karşılıklı ilişkide olduğu grubu oluşturmuşlardır.[52] Dinozorlar Çağı'nda üç büyük damarlı bitki grubu mevcuttu. Bunlar: tohumsuz bitkiler, açık tohumlular ve Kretase içinde evrimleşen kapalı tohumlu bitkiler. Eğrelti otu, kurt ayağı gibi bitkiler tohumsuz ürerler; hızlı büyüme ve hızlı üreme özellikleriyle nemli bölgelerde otçul dinozorların otlama potansiyellerini dengelemiştir. Daha sonra ilk tohumlu bitkilerin üyeleri olan açık tohumlu bitkiler evrimleşmiş ve birtakım modifikasyonları kazanır hâle gelmişlerdir, zira otsu ve tohumsuz bitkilere nazaran yavaş büyüyen ve üreyen açık tohumlularsa otçul baskısına daha açıktılar. Bu duruma karşı güçlü evrimsel cevaplar verildi, örnek olarak güçlü hücre duvarları, reçine üretimi, dikensi doku ve kalın ağaç gövdesi yapısı otçulların onları daha az tercih etmesini sağladı. Tohum avantajı uzak yerlere taşınma metoduyla dağılımı sağlayıp bitkilerin çevreye olan hâkimiyetinin önünü açtı. Kretase içinde kapalı tohumlu bitkiler de ortaya çıktı,[53] bir koruyucu doku yani meyve içinde kalan tohum daha iyi korunuyordu ve bu bitkiler açık tohumlu bitkilerden daha hızlı büyüyor ve ürüyorlardı. Bu nedenledir ki, günümüzde de bitkisel yaşamın en büyük üyeleri kapalı tohumlu bitkilerdir.[30]
Habitat
Dinozorlar Mezozoyik'te farklı kıtalarda çeşitli habitatlarda varlıklarını sürdürmüşlerdir. Kurak ortamlardan çöllere, taşkın ovalarından lagünlere ve orman ekosistemlerine dek farklı yaşam alanlarına dağılmışlardır. Geç Jura'da yaşayan Archaeopteryx'in fosilleri lagün ortamında bulunmuştur ki o dönem Avrupa'nın takım adalar halinde olduğu bilinir.[54]
Geç Kretase'de bugünkü Sahra çölü bir ırmak ekosistemiydi ve Spinosaurus ve Carcharodontosaurus menderes yapan ırmakların, taşkın ovalarının yer aldığı bu ortamda yaşamaktaydı.[55] Son yıllarda Spinosaurus'un özelinde spinosaur ailesinin üyelerinin yaşam biçimleri üzerinde çok fazla tartışma yapıldı. Kimi çalışmalarda su yüzeyinde yüzen veya su kaynaklarının kıyılarında gezen yarı sucul bir canlı (bozayı, kaz) gibi betimlenirken; bazı çalışmalarda bilim insanları tarafından su altında hareket eden, suya tamamen dalan ilk dinozor olarak düşünüldü.[56] Nathan P. Myhrvold ve ark. 2024 yılında yaptıkları çalışmada, makine öğrenmesine dayalı bir yöntem (pFDA) kullanarak Spinosaurus'un tamamen sucul yaşam biçimine sahip olduğunu ileri süren bir önceki çalışmanın verileri ve onların nasıl kullanıldığını yeniden değerlendirdi. Sonuç olarak yaratığın tamamen suya dalan ve avını su içinde deniz aslanı gibi takip eden bir yaşam biçimine sahip olmadığı düşünüldü. Daha çok su kıyısında 2 metreyi aşmayan sığlıklarda dolaşarak pençeleri ve ağzıyla balık yakalayabilecek bir yaşam biçimine sahip olduğu iddia edildi.[57]
Yine geç Kretase yaşlı ünlü Kuzey Amerika fosil faunasına sahip Hell Creek formasyonu da sulak kıyı ovaları ve menderes çizen ırmakların taşkın ovaları çökellerini barındırır. T-rex, Triceratops ve Edmontosaurus gibi dinozorlar bu nemli arkaik çevrede yaşadı.[58]
Güney Amerika dinozor fosili barındıran çokça jeolojik oluşuma sahiptir. Ünlü fosilli oluşumlardan biri de Geç Kretase yaşlı Candeleros formasyonudur. Giganotosaurus, Buitreraptor, Jakapil ve pek çok sürüngen bu formasyondan bilinmektedir. Bu formasyon, akarsu ortamı ve eoliyen ortamı yansıtan orta-büyük taneli kumtaşlarından oluşur. Silttaşı ve çamurtaşı varlığı da paleoçevrenin bataklık koşullarını temsil eder.[59]
Çin de zengin fosil içeriğine sahip formasyonlara sahiptir. Bunlardan biri Liaoning eyaletindeki Erken Kretase yaşlı Yixian formasyonudur. Buradaki kaya oluşumları periyodik olarak gerçekleşen volkanik patlamaların çökel kayıtlarındaki izlerini taşır ve bu nedenle Mezozoyik Pompeii olarak da adlandırıldı. Formasyonun temsil ettiği arkaik çevre yüksek bir canlı çeşitliliğiyle sıcak bir iklimi işaret eden ve çok sayıda gölü de barındıran bir çevreydi ve Dilong paradoxus gibi erken tyrannosauridlerden birini içeriyordu.[60]
Evrimsel süreçler
Dinozorlar, Mezozoyik boyunca arkaik yeryüzünde her kıtaya yayıldılar. Bu yayılış tüm türlerin yerküre boyunca sınırsız hareket ettiği bir dağılış değil, her türün kendine özgü bir dağılım şekline sahip olmasıyla tanımlanır. Bu yayılış büyük oranda kıtaların fiziksel konumları, iklim ve sıcaklık kuşaklarıyla ilgilidir.[50] Dinozorların çevreye muazzam ölçüde iyi adapte olması ve ekolojik anlamda tüm nişleri doldurması, kayda değer bir başarıdır. Bitkiler bile bu karşıtlık üzerinden kendi evrimsel rotalarını çizmişlerdir; örneğin Araucaria cinsi ağaçlar uzun boyunlu sauropodlardan korunmak adına alt dallarından kurtulmuşlardır. Yine Stegosaurus cinsi geç Jura dinozoru temel diyeti olan çikas türevi bitkilerin yok olmasıyla ortadan kaybolduğu ileri sürüldü. Dinozorların uzun evrimsel öyküsü ise aslında soylarının tamamen tükenmemesidir, bugün artık pek çok bilimsel çalışmada kuşlar dinozorların yaşayan torunları olarak kabul edilmektedir. Dinozorlarla kuşların aralarındaki akrabalık ilişkisini vurgulayan ilk bilim insanı Thomas Huxley'dir.[61] Uzun bir süre ilk kuş olarak adlandırılan Archaeopteryx lithographica'nın bulunmasıyla fosildeki korunmuş olan tüyler yıllar sonra ciddi bir tartışmaya yol açtı ve yıllar yılı soğukkanlı, pullu ve hantal sürüngenler olarak düşünülen dinozorların kuşlarla yakın akraba olabilecekleri üzerine araştırmalar yapıldı. Yakın zamanlarda Çin'de pek çok tüylü dinozor bulunması, kuşların kanatlarındaki tüy formasyonuna benzer olarak dinozorların kol kemiklerinde tüy kökü çıkıntıları bulunması önemli kanıtlardan biridir. Kuşlarla yakın akraba oldukları genetik olarak belli olan dinozorların ön kollarındaki parmak yapılarının kuşlara benzemesi ve birçok dinozorun kuşlar gibi sıcakkanlı olduklarının yaşadıkları bölge ve fosiller yardımıyla saptanması sonucu böylesi bir akrabalık bilimsel anlamda netleşmiştir.[18]
Dinozorların irdelenen bir diğer yönü de kimi grupların evrimsel olarak yakınsak özellikler göstermesidir. Örnek olarak tyrannosauridler ve abelisauridler büyük bedenlerine ve kafataslarına göre çok küçük kalan ön kollara sahiptirler. Teropodlar içinde bu küçük kollu olma eğilimine bir grup daha sahiptir: Carcharodontosauridler. 2022 yılında yapılan bir çalışmada Arjantin'de Huincul formasyonundan tanımlanan Meraxes gigas'ın fosil örneği tam bir ön kolu korumuştu ve bu da canlının büyük bedenine göre bir hayli ufak kalmaktadır. Sonuç olarak üç teropod grubunun da büyük bedenlere karşın küçük kollar barındırarak yakınsak evrim örneği sunduğu belgelendi.[62]
Ayrıca bilim insanlarının dinozorların nasıl göründüğüne ilişkin algısı bilimsel çalışmalar ışığında ciddi oranda değişmiştir. Hantal, kertenkele benzeri pullu canlılar olarak gösterilirken; bugün artık fosil kanıtlar sayesinde pek çok teropodun kuş akrabalarınınkine benzer tüy türevi yapılara sahip olduğunu biliyoruz. Özellikle bilim insanlarının bu konudaki anatomik görüşü de evrim geçirmiştir. En bilinen dinozor unvanını hâlâ sürdüren karizmatik teropod türü Tyrannosaurus rex ortalama olarak 12 metre uzunluğunda, 5 metre yüksekliğinde ve kabaca 9 ton ağırlığında[63] bir canlıydı, bu tahminler yaklaşık bir asır önceki keşfinden bu yana çok değişmedi; ancak bu canlının rekonstrüksiyonu oldukça değişmiştir. Tyrannosaurus hatta neredeyse tüm teropodlar için eski görüşler, kuyruğunu sürüyen, neredeyse insan gibi dik duran ve hantal bir varlık düşünürken; modern görüş hızlı, tüylü bir vücut ve eğik bir duruş betimlemiştir.[64]


Kuşlarda yaygın olarak gözlenen uyku pozisyonunun bazı dinozorlarda da görüldüğünü ortaya koyan fosil kalıntılar bulundu. 2004 yılında Xu Xing ve Mark Norell bir Troodontid olan Mei long'u tanımladı, bu dinozorun kalıntıları günümüz kuşlarında görülen uyku pozisyonunu yansıtan bir duruşla korunduğu anlaşıldı ve hayvanın uyurken aniden gelişen bir afetle gömüldüğü sanılıyor, fosil alanındaki tüf katmanları da volkanik bir etkinliği doğruladı.[65]

Tüylülük
Tüyler dinozorların bir grubu olan kuşların en göze çarpan özelliklerinden biridir. Son 30 yılda yapılan çalışmalarda ise kuş olmayan dinozorların çeşitli üyelerinin fosillerinde de tüy vb. yapıların kalıntılarına rastlandı ki bazı bilim insanlarına göre dinozor soy ağacının farklı gruplarında tüylü üyelerin bulunması tüylülüğün atasal bir özellik olduğu düşüncesini doğurdu.[66] Son yıllarda Çin'de bulunan tüylü kuş olmayan dinozorlardan sonra Tyrannosaurus rex gibi büyük Geç Kretase Tyrannosauridlerinin de tüylü olabileceği tartışıldı. Yakın zamana kadar tüylülüğün daha çok küçük dinozorlarda görülen özellik olduğu düşünüldü çünkü canlılar büyüdükçe yüzey alanı/hacim oranı düşer ve bu da ısı kaybını azaltır ve sonuç olarak beden sıcaklığını korumak için tüylere gereksinim kalmaz. Yutyrannus'un keşfiyle büyük bir teropodun da tüylü olabileceği görüldü ve büyük Geç Kretase tyrannosauridleri de tüylerle betimlenmeye başladı,[67] ancak Montana'da Tyrannosaurus rex'e ait olan fosil bir deri baskısı bulunmasıyla hayvanın pullarla kaplı olduğu anlaşıldı. Büyük Tyrannosauridlerin tüy barındırsalar bile bunların gövdelerinin yanlarıyla sınırlı olduğu düşünüldü.[68]
Erken Kretase döneminde bugünkü Çin'de yaşayan Psittacosaurus'un da kuyruğunun gerisinde tüy benzeri yapılar taşıdığı çok iyi korunmuş bir fosil parçası sayesinde belirlendi. Tüy benzeri yapıların gösteriş veya uyarı için kullanılmış olabileceği ileri sürüldü. Bu yapıların tam olarak tüylerle mi yoksa epidermal çıkıntılar mı homolog olduğu hala tartışmalı olsa da teropod olmayan bir dinozorda tüy benzeri bir yapının rapor edilmesi önemliydi.[69]
Sert doku
Dinozorların bazı grupları fiziksel görünüm konusunda sıradışı olan canlıları barındırır. Boynuzlar, dikenler, plakalar, topuzlar ve yumrular gibi özelleşmiş yapılar dinozorların ekosistem içindeki evrimsel gelişiminin bir sonucudur. Bu yapıların işlevi hakkında 5 temel açıklama ileri sürüldü: Fizyolojik görevler, savunma,[70] cinsel seçilim, tür içi rekabet ve tür içi tanınma.[71] En ünlü dinozorlardan biri olan Stegosaurus'un karakteristik görünümünü borçlu olduğu sırt plakaları da işlevlerinin özelinde çok tartışıldı. Başlangıçta savunma amaçlı olduğu düşünülen bu fosil yapıların görece kırılgan ve damarlı iç yapıları nedeniyle termoregülasyon işlevi gördüğü ileri sürüldü.[72] Bir avcıya karşı gösterilen savunma davranışına ilişkin fosil kanıt bir Allosaurus bireyinin pelvisine saplanmış Stegosaurus kuyruk dikenidir.[73] Sadece otçul dinozorlarda değil teropodlarda da boynuz ve kemiksi çıkıntılar görülür. Boynuza sahip sahip tek teropod Carnotaurus'tur ve bu boynuzların tür içi rekabet veya çiftleşme döneminde dişileri etkilemekte kullanıldığı öne sürüldü.[74] Özelleşmiş yapılara sahip dinozorlar içinde olan bir diğer grup da ankylosaurlardır. Zırhlı dinozorlar olarak bilinen bu grubun en yi bilinen cinsi Ankylosaurus'tur. Bu cins, sırtındaki kemiksi plakaların yanı sıra kuyruğunun ucunda yırtıcılara veya tür içi rakiplerine karşı savurduğu bir topuz da bulundurur.[75]
Yumuşak doku
Yumuşak dokular, fosilleşmeleri çok zor olduğu için seyrek olarak fosil kayıtlarda bulunurlar. Yine de dinozor çalışmaları içinde önemli yumuşak doku keşifleri yapıldı. Daha 19.yy'da pullu deri örnekleri bulundu, ama bunlar o dönemde yanlış olarak timsahlara atfedildi. İlgili bulgu, 20. yy'ın ilk çeyreğinde Reginald Hooley tarafından daha ayrıntılı olarak incelendi.[76] Bunun yanı sıra sonradan Edmontosaurus olarak bilinen dinozorun kalıntıları arasında da yumuşak dokulara rastlandı ve 1912'de H. F. Osborn tarafından pullu derili dinozor betimlendi.[77]
Teropodlar arasında da yumuşak dokuları korumuş fosillere sahip dinozorlar bulundu. İtalya'da bulunan ve Scipionyx adı verilen dinozor çarpıcı biçimde kas yapılarını koruyan ve iç organ yerleşimlerini gösteren kalıntılara sahipti.[78]
Yakın tarihli çalışmalar da önemli bulguları ortaya koydu. 2022 yılında Bell ve ark. Psittacosaurus fosili üstünde pul yapılarına rastladı. Hayvanın kuyruğunda dörtgenel pullar varken, omuzlarındakiler kesik koni biçimindeydi.[79] Ayrıca Dakota lakabı verilen Edmontosaurus fosili çok iyi korunmuş bir ön ayak örneğine sahipti, canlının ön ayaklarındaki orta parmakların birleşerek günümüz atlarındaki toynaklara benzeyen bir yapıyı oluşturduğu bulundu.[80]
Büyük grupların yok oluşu
Büyük dinozorların nasıl yok olduğuna dair bugüne değin birçok iddia ortaya atılmıştır. Geçmişte, dinozorların kısa bir süre içinde toplu olarak nasıl yok oldukları uzun bir süre açıklanamamış ve yanardağ patlamalarından dünyadaki iklim değişikliklerine kadar çeşitli teoriler ortaya atılmıştır. Bu konuda en yeni teori gök taşı (asteroid) çarpması sonucu bir felaket zinciri hâlinde yok oluşun tetiklendiği varsayımıdır. Yaklaşık 66 milyon yıl önceye tarihlenen yani stratigrafik olarak Kretase-Paleojen yok oluşu dizisi diye adlandırılabilecek bir çökel istif bu yok oluşu temsil ediyor.[81]
Ünlü jeolog Walter Alvarez İtalya'nın Gubbio şehrinde bu yok oluş anını aydınlatacak önemli bir jeolojik iz keşfetti; bu Kretase döneminin sonunu temsil eden kil tabakasındaki anomaliydi. Bu anomali K-T sınırındaydı ve fosilsizdi. Bu jeolojik sınır Kretase ve Paleojen yaşlı birimlerin oluşturduğu tezatları gösteriyordu zira fosilce zengin Kretase kireçtaşları, ince bir kil tabakası ardından fosilsiz Paleojen kayaçlarına yerini bırakıyordu. Bu noktada kil tabakası jeolojik tarihte nasıl bir olayı betimliyordu sorusu gündeme gelmiştir ve bu sorunsalın yanıtına Walter'ın babası olan ünlü fizikçi Luis Alvarez 1970'lerde Gubbio şehrinde bu jeolojik sınırda nadir bir element olan iridyumu bularak yaklaşmıştır.[82] Dünya'nın kabuğunda milyonda 0,3 olarak bulunan bu element (sondaj karotları veyahut yüzlek veren kayalardan örnekler toplanarak), K-T yok oluş sınırında milyonda 6,3'lük bir oranda bulundu yani 20 kat daha fazla bir miktar vardı. Dolayısıyla bugün Güneş sistemi bünyesindeki pek çok asteroidin de ortalama olarak bu düzeyde bir iridyum varlığına sahip olduğu bilindiğinden yaklaşık 66 milyon yıl önce Dünya'nın yüzeyine çarpan dev bir asteroidin toplu yok oluşun bir dizi sebebini ortaya çıkarabileceği mantıklı bir görüş hâline gelmiştir. Çapı 10 km,[83] ağırlığı ise 1 katrilyon ton olarak hesaplanan bu dev asteroidin çarptığı yerin Meksika'daki Yucatan yarımadası Chicxulub krateri olduğu tahmin ediliyor. Çarpışma anında hızının saatte 100 bin km olabileceği düşünülürken, 11.0 Mw büyüklüğünün[84] üstünde bir depremi tetiklemiştir (beraberinde dev tsunamiler) ve sonuç olarak böylesine bir çarpışma alanı (astrobleme) yüzlerce km çapında bir alanı kaplayabilir.
Daha da ilginci son araştırmalarda Ukrayna Boltiş kraterinin Chicxulub krateri ile hemen hemen aynı yaşta olduğu Ukrayna'daki kraterin sadece 2000-5000 yıl daha yaşlı olduğu görülmüştür. Bu durum da paleontologlara K-T yok oluşunun çoklu bir dünya dışı etkinin sonucu olduğunu düşündürmektedir. Kafa karıştıran bir nokta da dünyaya nazaran oldukça küçük bir asteroidin taşıdığı iridyum nasıl olur da K-T stratigrafik sınırı boyunca tüm yerküreye görece istikrarlı dağılabilmiştir; ama bu soruya rağmen asteroid hipotezi yaygın kabul görür. Bu noktada asteroid çarpması, dinozorları yeryüzünden anında sildi açıklaması doğru değildir, bu teorinin devamına kulak vermek gerekir. Çarpışma sonucu büyük miktarda kıtasal ve meteorik parça atmosfere saçıldı ve sonuçta devasa bir toz-partikül (silikat parçaları) bulutu dünyayı sararak güneş ışığının yeryüzüne ulaşmasını engelledi. Bu ise zincirleme birtakım felaketlere neden oldu, zira önce fotosentetik canlıların soyları tükendi (büyük oranda bitkilerin) ve bu da herbivor (otçul) dinozorların kademe kademe yok oluşuna gitti ve nihayet carnivor (etçil) dinozorlar bu yok oluş silsilesinden etkilendi.[18][85]
Yukarıdaki söylendiği gibi günümüzde kuş olmayan dinozorları yok eden olayın gerçekleşme şekli hâlâ tartışma konusudur. Kuş olmayan dinozorların yok oluşuna ilişkin en önemli tartışma konularından birisi de bu yok oluşun ani (katastrofik) mi, yoksa aşamalı olarak mı gerçekleştiğidir.[86] Çin'de dinozor yumurtaları üzerinde yapılan bazı çalışmalara göre Kretase-Tersiyer yok oluşu öncesi iki milyon yıllık bir zaman diliminde yaklaşık 1000 kadar yumurta kabuğu ve tam yumurta fosilinin sadece üç farklı yumurta kabuğu çeşidinden geldiği belirlenmiştir. Bu üç çeşit yumurta kabuğundan ikisi dişsiz oviraptorlardan gelirken diğeri otçul hadrosaurlarınkidir. Bu veriler ışığında yok oluş öncesinde zaten dinozorların çeşitlilik düşüşüne uğradığı ve yok oluşun aşamalı olduğu ileri sürülebilir.[87]
Genelde fosil çalışmaları gösteriyor ki, toplu yok oluşlar ufak türleri es geçmektedir. Bilhassa özel nişleri dolduran ve belli bir oranın üstünde vücut büyüklüğü bulunan canlılar yok oluşları atlatamamıştır. K-T yok oluşu da kuşlar, sürüngenler ve memelilerin atlattığı; ama kuş olmayan dinozorların atlatamadığı bir dönemeç olarak görülebilir. Son olarak belirtmek gerekir ki, Kretase periyodunun sonuna doğru Dekkan yaylasındaki volkanik kapanların[88] da püskürdüğü ve onların da bu yok oluşta bir paylarının olduğu düşünülebilir.[18][89]
Yakın zamanda yapılan bir çalışmada kuş olmayan tüm dinozorları ve diğer bazı canlıları yok eden Geç Kretase sonu toplu yok oluş olayının Kuzey yarımküre'nin ilkbahar aylarında yaşandığı belirlenmiştir. Bu çalışmada yok oluşun mevsimi Acipenseriform grubu balıkların kalıntılarındaki duraylı izotoplardan yola çıkılarak mevsimsel bir döngü üzerinden bulunmuştur.[90]
Olası Paleosen dinozorları
Kimi zaman Kretase-Paleosen yok oluşu sınırının üstünde kuş olmayan dinozor kalıntılarına rastlanır. Bu kalıntıların bazılarının aşınma sonucu Geç Kretase yaşlı katmanlardan kopup Paleosen yaşlı kayalarda yeniden gömüldüğü anlaşıldı. Ancak Fasset ve ark. (2011), New Mexico eyaletindeki Ojo Alamo Sandstone oluşumundan çıkan bir Alamosaurus sol uyluk kemiğini (kemikten boyuna ve enine enkesitler alarak) Uranyum-kurşun dönüşümü kullanarak doğrudan yaşlandırdı ve bu fosil örneğin yaşı 63.4 ± 0.6 milyon yıl önceye tarihlendirildi.[91]
Fassett ve ark. (2009, 2011)'nın ortaya attığı Paleosen dinozorları hipotezine bazı araştırmacılar tarafından itiraz edildi. Lucas ve ark. (2011)'de Fassett'in çıkarımlarına stratigrafik yönlerde karşı çıktı, ilk olarak Ojo Alamo kumtaşının Kretase-Paleosen yok oluş çizgisini içerdiğini ve alt Ojo Alamo kumtaşının yaşının Maastrichtian olduğunu iddia ettiler. Bu hipoteze getirilen önemli bir itiraz da Paleosen çökellerine bulunan hadrosaur bacak kemiğinin Geç Kretase yaşlı tabakalardan taşındığı ve Paleosen tabakalarına yeniden çökeldiği ve dolayısıyla Paleosen'de kuş olmayan dinozorların yaşamadığının öne sürülmesiydi.[92] Kalıntıların parçalı ve aşınmış olması kanıt olarak sunuldu. Paleosen dinozorları hipotezine bir diğer karşı çıkış da Koenig ve ark. (2012)'nın yaptığı çalışmada dile getirildi, bu çalışma özetle Fassett'in 2011 tarihli çalışmasında yaptığı radyoaktif yaşlandırma ile çok geniş bir yaş aralığı bulduğunu, hipotezi için yeterli paleopalinolojik ve manyetostratigrafik kanıtlar öne süremediğini ve yaşlandırma noktasında yeniden kristallenme işleminin süresini belirlemediğini savundu.[93]
Bu konudaki bir diğer görüş de Kretase-Paleosen yok oluş çizgisinin 1.3 metre üstünde bulunan ve Paleosen dönemine tarihlenen çökellerde yer alan dinozor dişlerine dayanarak kuş olmayan dinozorların yok oluştan sonra yaklaşık 40.000 yıl boyunca yaşadığı savıdır. Özellikle dişler üstünde aşınma ve deformasyon olmaması da bu dişlerin daha yaşlı tabakalardan Paleosen çökellerine taşındığı iddiasını çürütebilir. Sonuç olarak, olasılıkla Kretase-Paleosen yok oluşundan binlerce yıl (yaklaşık 40 bin yıl) sonra bile yerküre üzerinde kuş olmayan dinozorların yaşadığı düşünülüyor.[94]
Ayrıca bakınız
Notlar
- ^ Dinozor adı, Türkçeye Fransızcadaki dinosaur teriminden geçmiştir.
Kaynakça
- Özel
- ^ Matthew G. Baron; Megan E. Williams (2018). "A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution". Acta Palaeontologica Polonica. 63. DOI:10.4202/app.00372.2017.
- ^ Andrea Cau (2018). "The assembly of the avian body plan: a 160-million-year long process" (PDF). Bollettino della Società Paleontologica Italiana. 57 (1): 1–25. DOI:10.4435/BSPI.2018.01.
- ^ Ferigolo, Jorge; Langer, Max C. (1 Ocak 2007). "A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone". Historical Biology. 19 (1): 23-33. doi:10.1080/08912960600845767. ISSN 0891-2963.
- ^ Langer, Max C.; Ferigolo, Jorge (1 Ocak 2013). "The Late Triassic dinosauromorph Sacisaurus agudoensis (Caturrita Formation; Rio Grande do Sul, Brazil): anatomy and affinities". Geological Society, London, Special Publications (İngilizce). 379 (1): 353-392. Bibcode:2013GSLSP.379..353L. doi:10.1144/SP379.16. ISSN 0305-8719. 14 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 1 Temmuz 2023.
- ^ Cabreira, S.F.; Kellner, A.W.A.; Dias-da-Silva, S.; da Silva, L.R.; Bronzati, M.; de Almeida Marsola, J.C.; Müller, R.T.; de Souza Bittencourt, J.; Batista, B.J.; Raugust, T.; Carrilho, R.; Brodt, A.; Langer, M.C. (2016). "A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet". Current Biology. 26 (22): 3090-3095. doi:10.1016/j.cub.2016.09.040
 . ISSN 0960-9822. PMID 27839975.
. ISSN 0960-9822. PMID 27839975. - ^ Müller, Rodrigo Temp; Garcia, Maurício Silva (26 Ağustos 2020). "A paraphyletic 'Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs". Biology Letters. 16 (8): 20200417. doi:10.1098/rsbl.2020.0417 . PMC 7480155 $2. PMID 32842895.
- ^ "etymology of Dinosaur word". 17 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Nisan 2024.
- ^ Sakamoto, Manabu; Benton, Michael J.; Venditti, Chris (3 Mayıs 2016). "Dinosaurs in decline tens of millions of years before their final extinction". Proceedings of the National Academy of Sciences (İngilizce). 113 (18): 5036-5040. doi:10.1073/pnas.1521478113. ISSN 0027-8424. PMC 4983840 $2. PMID 27092007. 15 Temmuz 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 15 Temmuz 2024.
- ^ a b Paul, Gregory. "DETERMINING THE LARGEST KNOWN LAND ANIMAL: A CRITICAL COMPARISON OF DIFFERING METHODS FOR RESTORING THE VOLUME AND MASS OF EXTINCT ANIMALS" (PDF). www.gspauldino.com. ANNALS OF CARNEGIE MUSEUM. 28 Ocak 2020 tarihinde kaynağından (PDF) arşivlendi.
- ^ "The functional origin of dinosaur bipedalism: Cumulative evidence from bipedally inclined reptiles and disinclined mammals". 27 Ocak 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Ocak 2024.
- ^ Brownstein, Chase D. "The biogeography and ecology of the Cretaceous non-avian dinosaurs of Appalachia" (PDF). Palaeontologia Electronica. 20 Temmuz 2018 tarihinde kaynağından (PDF) arşivlendi.
- ^ The complete dinosaur
- ^ Editors, Editors. "The truth about Griffins..." www.liverpoolmuseums.org.uk. 11 Ağustos 2020 tarihinde kaynağından arşivlendi.
- ^ "Did the horned dinosaur Protoceratops inspire the griffin?". 24 Haziran 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Haziran 2024.
- ^ James (Çev. Cumhur Öztürk), Ioan (2009). Büyük Biyologlar Ray'den Hamilton'a (2021 bas.). Cambridge: Türkiye İş Bankası Kültür Yayınları. s. 129. ISBN 978-625-405-266-8.
- ^ Clark, Neil; Lindsay, William (1995). Pocket Dinosaurs. New York: Dorling Kindersly. s. 138. ISBN 1-56458-662-6.
- ^ Editors, Editors. "Iguanodon". DKfindout. 22 Ağustos 2016 tarihinde kaynağından arşivlendi.
- ^ a b c d e f Padian and Ostrom, Kevin and John. "Dinosaur". Encyclopædia Britannica. Encyclopædia Britannica, inc. 10 Haziran 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 20 Ocak 2020.
- ^ "Bone Wars". 18 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Nisan 2024.
- ^ Hellman, Hal (2001). Bilim Tarihinden Seçilmiş On Tartışma Büyük Çekişmeler. İstanbul: Tübitak. ss. 135-157. ISBN 975-403-211-4.
- ^ Peczkis, Jan. "Implications of body-mass estimates for dinosaurs". 24 Eylül 2015 tarihinde kaynağından arşivlendi.
- ^ a b Richardson, Hazel (2003). Dinosaurs and Prehistoric Life. Londra: Dorling Kindersly. ss. 68-69. ISBN 9780751337341. Erişim tarihi: 5 Temmuz 2023.
- ^ Persons IV, W. Scott; Currie, Philip J; Erickson, Gregory M. "An Older and Exceptionally Large Adult Specimen of Tyrannosaurus rex". 3 Ekim 2019 tarihinde kaynağından arşivlendi.
- ^ a b Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs. New Jersey: Princeton University Press. ss. 50-125-200. ISBN 978-0-691-16766-4.
- ^ Dalman, Sebastian G.; ve diğerleri. "A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism". www.nature.com. Nature. 11 Ocak 2024 tarihinde kaynağından arşivlendi.
- ^ [Re-assessment of the Late Jurassic eusauropod Mamenchisaurus sinocanadorum Russell and Zheng, 1993, and the evolution of exceptionally long necks in mamenchisaurids/ The longest neck]
- ^ "Global latitudinal gradients and the evolution of body size in dinosaurs and mammals". 11 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Nisan 2024.
- ^ Greshko, Michael. "DINOSAURS PREHISTORIC ICONS GET A MODERN REBOO". www.nationalgeographic.com. Nat Geo. 18 Şubat 2021 tarihinde kaynağından arşivlendi.
- ^ Phillips, Louis (1997). Ask me anything about dinosaurs. New York: Avon Books. ss. 32. ISBN 0-380-78552-8.
- ^ a b Surman, Holtz Jr., Farlow and Walters, M. K. Brett, Thomas R., James and Bob (2012). The complete dinosaur. Indiana, USA: Indiana University Press. ss. 650-660. ISBN 978-0-253-00849-7.
- ^ "Growth rates". 4 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Nisan 2024.
- ^ Osteohistological insight into the growth dynamics of early dinosaurs and their contemporaries[]
- ^ "Oviraptor is a parent, not a thief". 11 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Mayıs 2024.
- ^ a b Dixon, Dougal (2008). World Encyclopedia of Dinosaurs & Prehistoric Creatures. Lorenz Books. s. 31. ISBN 978-0754817307.
- ^ a b c Currie, Philip; Padian, Kevin (1997). Encyclopedia of DINOSAURS. Amsterdam: Academic Press. ISBN 978-0122268106.
- ^ "hadrosaurs lived in herds". 11 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Mayıs 2024.
- ^ [Spielberg was right: Triceratops teamed up/ Did Triceratops live together?]
- ^ Tanke, Darren H.; Currie, Philip J. "Head-biting behavior in theropod dinosaurs: Paleopathological evidence" (PDF). www.researchgate.com. GAIA - Ecological Perspectives for Science and Society. 27 Şubat 2008 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 14 Ocak 2024.
- ^ Drumheller, Stephanie K. "High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem". journals.plos.org/. PLOS ONE. 31 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Ocak 2024.
- ^ ""The Fighting Dinosaurs": The position of their bodies before and after death Published: 11 February 2017". 10 Kasım 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2024.
- ^ "EVIDENCE OF PREDATORY BEHAVIOR BY CARNIVOROUS DINOSAURS". 25 Şubat 2024 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 3 Mayıs 2024.
- ^ "Irritator lower jaw". 27 Mart 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mart 2024.
- ^ "Irritator anatomy". 11 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Mayıs 2024.
- ^ de Lazaro, Enrico. "Eggs of Earliest Dinosaurs Were Leathery, Fossils Show". www.sci.news/. SciNews. 14 Kasım 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 13 Kasım 2023.
- ^ Timperley, Jocelyn. "Scientists have discovered the tell-tale signs of a range of dinosaur diseases – and found that they're remarkably similar to those affecting animals alive today". BBC. 8 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 8 Nisan 2023.
- ^ respiratory disease in the first known non-avian dinosaur 16 Ocak 2024 tarihinde Wayback Machine sitesinde arşivlendi..
- ^ Black, Riley. "First dinosaur found with a respiratory disease". www.natgeo.com. National Geographic. 17 Mayıs 2022 tarihinde kaynağından arşivlendi.
- ^ "New information on paleopathologies in non-avian theropod dinosaurs: a case study on South American abelisaurids". 28 Şubat 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Nisan 2024.
- ^ Editors, Editors. "The man who split the dinosaurs in two". Geological Society of London. 25 Mart 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Temmuz 2023.
- ^ a b c Brusatte, Stephen L. (2012). Dinosaur Paleobiology. West Sussex: Wiley-Blackwell. ss. 20-22. ISBN 978-0-470-65657-0.
- ^ Dinosaurs and land plants.
- ^ Black, Riley. "Flowers, Pine Cones and Dinosaurs". Smithsonianmag. Smithsonian Enstitüsü. 24 Ocak 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 5 Temmuz 2023.
- ^ Angiosperms and dinosaurs
- ^ Ksepka, Daniel T. "Flights of Fancy in Avian Evolution". www.americanscientist.org. American Scientist. 7 Mayıs 2018 tarihinde kaynağından arşivlendi.
- ^ Baggaley, Kate. "The Sahara Desert was once flooded with history's most vicious dinosaurs". www.popsci.com. Popular Science. 1 Mayıs 2020 tarihinde kaynağından arşivlendi.
- ^ "Subaqueous foraging among carnivorous dinosaurs". 11 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mart 2024.
- ^ "Paleontologists Find Flaws in Research Supporting Aquatic Pursuit Hunting in Spinosaurus". 29 Mart 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mart 2024.
- ^ Davide, Fastovsky; Antoine, Bertovici. "The Hell Creek Formation and its contribution to the Cretaceous– Paleogene Extinction: a short primer" (PDF). www.sciencedirect.com. Sciencedirect.
- ^ Leanza, Héctor A.; Apesteguia, Sebastian; Novas, Fernando E.; de la Fuente, Marcelo S. "Cretaceous terrestrial beds from the Neuquén Basin (Argentina) and their tetrapod assemblages". www.sciencedirect.com. Cretaceous Research. 12 Haziran 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 20 Eylül 2023.
- ^ "Liaoning Province—China's Extraordinary Fossil Site". 24 Haziran 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Haziran 2024.
- ^ Huxley's thoughts
- ^ Canale, J. I.; ve diğerleri. "New giant carnivorous dinosaur reveals convergent evolutionary trends in theropod arm reduction". pubmed.ncbi.nlm.nih.gov/. National Library of Medicine. 18 Ekim 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 13 Ocak 2024.
- ^ Therrien and Henderson, François and Donald. "My theropod is bigger than yours...or not: Estimating body size from skull length in theropods is bigger than yours...or not: Estimating body size from skull length in theropods" (PDF). dinoweb.ucoz.ru. DINOWEB. 2 Ekim 2013 tarihinde kaynağından (PDF) arşivlendi.
- ^ Ross & Haas & Allmon, R. & D. & W. "Drawing Tyrannosaurus – You're Probably Doing it Wrong". www.nationalgeographic.com. Nat Geo. 19 Mart 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Mart 2020.
- ^ "A new troodontid dinosaur from China with avian-like sleeping posture". 26 Ocak 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 26 Ocak 2024.
- ^ Switek, Brian. "Rise of the fuzzy dinosaurs". www.nature.com. Nature. 13 Nisan 2020 tarihinde kaynağından arşivlendi.
- ^ Xu, Xing; ve diğerleri. "A gigantic feathered dinosaur from the Lower Cretaceous of China". Nature. 29 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Nisan 2023.
- ^ Katz, Brigit. "T. Rex Was Likely Covered in Scales, Not Feathers". www.smithsonianmag.com. Smithsonian Magazine. 29 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Nisan 2023.
- ^ "Psittacosaurus and its bristle-like structures". 13 Mart 2024 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 14 Nisan 2024.
- ^ "Horns, Clubs, Plates and Spikes: How Did They Evolve?". 3 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2024.
- ^ "The evolution of 'bizarre structures' in dinosaurs: biomechanics, sexual selection, social selection or species recognition?". 16 Aralık 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2024.
- ^ "Plates of the dinosaur stegosaurus: forced convection heat loss fins?". 3 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2024.
- ^ "Spiked tail to the rescue!". 3 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2024.
- ^ "Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei". 3 Kasım 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2024.
- ^ "Ankylosaurs may have used their tail clubs to fight each other, not predators". 4 Nisan 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2024.
- ^ On the Integument of Iguanodon bernissartensis, Boulenger, and of Morosaurus becklesii, Mantell[]
- ^ 28b. Trachodon Twice Restored, 1912[]
- ^ "Scipionyx soft tissues". 30 Nisan 2024 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 11 Mayıs 2024.
- ^ "The exquisitely preserved integument of Psittacosaurus and the scaly skin of ceratopsian dinosaurs". 12 Ağustos 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Şubat 2024.
- ^ "Incredible dinosaur 'mummy' reveals a surprisingly hoof-like foot". 23 Aralık 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Şubat 2024.
- ^ Editors, Editors. "Kretase Yok Oluşu: Sürüngenler Çağının Sonu". www.tubitak.com. Tübitak. 11 Aralık 2005 tarihinde kaynağından arşivlendi.
- ^ Brusatte, Steve (2020). Dinozorların Yükselişi ve Çöküşü. İstanbul: Koç Üniversitesi Yayınları. ss. 210-215. ISBN 978-605-7685-46-9.
- ^ "The Chicxulub impactor". 11 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Mayıs 2024.
- ^ Schulte, Peter; ve diğerleri. "The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous-Paleogene Boundary" (PDF). www.sciencemag.com. Sciencemag. 21 Eylül 2017 tarihinde kaynağından (PDF) arşivlendi.
- ^ Senel, Cem Berk; ve diğerleri. "Chicxulub impact winter sustained by fine silicate dust". www.nature.com. Nature Geoscience. 20 Kasım 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Kasım 2023.
- ^ "The Great Mystery". 23 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Mayıs 2024.
- ^ Editor, Editor. "Dinosaur Biodiversity Was in Decline Millions of Years before End-Cretaceous Extinction: Study". www.sci.news. News Staff / Source. 27 Eylül 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Eylül 2022.
- ^ "DID 'VOLCANIC WINTERS' LEAD TO DINOSAUR EXTINCTION?". 26 Ocak 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 26 Ocak 2024.
- ^ "Deccan Traps". 11 Mayıs 2024 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Mayıs 2024.
- ^ During, Melanie A.D. "The Mesozoic terminated in boreal spring". www.nature.com. Nature. 24 Şubat 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Ağustos 2022.
- ^ Fassett, James E.; Heaman, Larry M.; Simonetti, Antonio. "Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico". 2 Aralık 2017 tarihinde kaynağından arşivlendi.
- ^ Lucas, S.; ve diğerleri. "No Definitive Evidence of Paleocene Dinosaurs in the San Juan Basin". Semantic Scholar. 30 Nisan 2023 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Nisan 2023.
- ^ Koenig, Alan E.; ve diğerleri. "Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico: COMMENT". www.usgs.com. USGS. 9 Mayıs 2020 tarihinde kaynağından arşivlendi.
- ^ SLOAN, ROBERT E.; RIGBY, J. KEITH; M. VAN VALEN, LEIGH; GABRIEL, DIANE. "Gradual Dinosaur Extinction and Simultaneous Ungulate Radiation in the Hell Creek Formation". Science. 4 Kasım 2021 tarihinde kaynağından arşivlendi.
- Genel
- Encyclopedia of dinosaurs & prehistoric life. American Museum of Natural History. New York, N.Y.: DK Publishing in association with the American Museum of Natural History. 2008. ISBN 978-0-7566-3836-8. OCLC 181602635.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




















