Yarasa
| Yarasalar | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Biyolojik sınıflandırma | |||||||||
| |||||||||
| Alt takımlar | |||||||||
(geleneksel):
(yeni):
| |||||||||
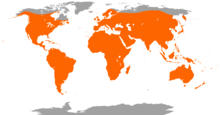 | |||||||||
| Yarasa türlerinin dünya üzerinde dağılımı | |||||||||
Yarasalar, ön ayakları kanat olarak uyarlanmış ve doğal olarak gerçekten uçabilen Chiroptera[a] takımında sınıflandırılan memelilerdir. Yarasalar, üzerleri zar ve patagium ile kaplı çok uzun ve yayılmış parmaklarıyla kuşlardan daha kolay manevra yaparak uçabilirler. En küçük yaşayan memeli olduğu da iddia edilen yabanarısı yarasası 29 ila 34 mm. boyunda, 15 cm. kanat açıklığına sahip ve 2 ila 2,6 gram ağırlığındadır. En büyük yarasalar da "uçan tilki" adı verilen Pteropus cinsi yarasalardır. Acerodon jubatus türü 1,6 kg ağırlığındadır ve kanat açıklığı 1,7 m'yi bulur.
Kemiricilerden sonra memelilerin ikinci en büyük takımı olan yarasalar 1.200'den fazla türü ile dünya üzerinde tüm sınıflandırılmış memelilerin yaklaşık %20'sini oluşturur. Yarasalar geleneksel olarak iki alt takıma ayrılırdı: Meyve yiyen büyük yarasalar (Megachiroptera) ve ekolokasyon kullanan küçük yarasalar (Microchiroptera). Ancak son zamanlarda bulunan –daha çok genetik– kanıtlar doğrultusunda takım Yinpterochiroptera ve Yangochiroptera olarak ikiye ayrılmıştır ve küçük yarasaların bazı türleri büyük yarasalar ile ilk takımda sınıflandırılmıştır. Yarasaların çoğu böcekçildir ve kalanların çoğu da meyvecildir. Birkaç tür böceklerden başka hayvanlarla beslenir; örneğin Yeni Dünya'ya özgü vampir yarasalar çeşitli memelilerin ve büyük kuşların kanını emerek beslenirler. Yarasaların çoğu gececildir ve çoğu mağaralarda ya da başka korunaklarda tünerler. Bu davranışın nedeninin avcılardan kaçmak olup olmadığı bilinmemektedir. Aşırı soğuk bölgeler dışında yarasalar tüm dünyaya yayılmışlardır. Çiçeklerin polenlerini taşıma ve tohumları yayma gibi görevlerle ekosistemleri içinde önemli yere sahiptirler ve çoğu tropikal bitki bu hizmetler için tamamen yarasalara bağımlıdır.
Yarasalar, tehdit oluşturmanın yanı sıra insanlara yararlı da olmaktadır. Yarasa dışkısı mağaralardan guano olarak toplanır ve gübre olarak kullanılır. Haşere böcekleri yiyen yarasalar pestisit gereksinimini azaltır. Bazen turistlerin görmek isteyeceği kadar çok sayıda olabilirler ve Asya ile Pasifik Kıyılarında besin olarak tüketilirler. Kuduz gibi birçok patojenin doğal konaklarıdır ve çok hareket ettikleri, sosyal hayvanlar oldukları ve uzun yaşadıkları için çok kolay hastalık yayabilirler. Birçok kültürde yarasalar genellikle karanlık, kötü niyet, cadılık ve büyücülük, vampirler ve ölümle bağdaştırılırlar.
Etimoloji
Dîvânü Lugati't-Türk'te "Aya Yersgü: Yarasa. Çiğilce. Bir takımları yarısa derler." diye geçer.[4] Tuncer Gülensoy'a göre "*yarı 'deri+sa'" kelimesinden türetilmiş olabilir.[5] Osmanlıcada Arapçadan geçen "huffâş" (خفاش) kelimesi kullanılır. Nişanyan Sözlük'e göre yarsıġu veya yarasa "yarasa" sözcüğünden evrilmiştir ve yarsı- "iğrenmek, tiksinmek" fiilinden Eski Türkçe +gU ekiyle türetilmiş olabilir ancak bu kesin değildir.[6]
Anatomi ve fizyoloji
Kafatası ve dişler

Kafa ve dişlerin şekli türlere göre değişiklik gösterir. Genel olarak büyük yarasaların burunları daha uzun, göz çukurları daha büyük ve kulakları daha küçük olur. Bu şekilde köpeğe benzer bir görünüşe sahip oldukları için de "uçan tilkiler" olarak anılırlar.[7] Küçük yarasalarda uzun burun nektar ile beslenmeyle bağlantılı iken[8] vampir yarasaların burnu daha büyük kesici ve köpek dişlerine yer açmak için daha küçüktür.[9]
Küçük böcekçil yarasaların 38 kadar dişi olurken vampir yarasaların 20 kadar dişi vardır. Kalın kabuklu böceklerle beslenen yarasaların daha az ama daha büyük dişleri olur; köpek dişleri uzundur ve daha yumuşak gövdeli böceklerle beslenen yarasalara göre alt çeneleri daha sağlamdır. Nektar ile beslenen yarasalarda ise köpek dişleri uzunken azı dişleri küçüktür. Meyve ile beslenen yarasalarda azı dişlerinin üstü ezmek için uyum sağlamıştır.[8] Bu beslenme davranışları hem büyük hem de küçük yarasalar için geçerlidir. Vampir yarasaların üst kesici dişlerinde mine olmaması sayesinde jilet keskinliğindedirler.[9] Küçük yarasaların ısırma kuvveti kuvvet kazancı sayesinde oluşur ve bu şekilde böceklerin sertleşmiş kabuklarını ve meyvelerin kabuklarını ısırabilirler.[10]
Kanatlar ve uçuş

Yarasalar, süzülerek uçan uçan sincapların aksine gerçekten uçma yeteneğine sahip tek memelilerdir.[11] En hızlı uçabilen yarasa olan Tadarida brasiliensis 160 km/h yer süratine ulaşabilir.[12]
Yarasaların parmak kemikleri düzleşmiş kesitleri ve uçlara doğru düşük kalsiyum düzeyleri sayesinde diğer memelilere göre çok daha esnektir. Kanat gelişimi için gerekli bir özellik olan parmak kemiklerinin uzaması kemik morfojenik proteinlerinin (Bmp) artışı sayesindedir. Embriyo gelişimi sırasında Bmp işaretlemesini kontrol eden gen Bmp-2 yarasaların önayaklarında artmış ifadeye maruz kalır ve sonucunda da parmak kemikleri uzar. Bu kritik genetik değişiklik uçmak için özelleşmiş uzuvların oluşmasına yardımcı olur. Yaşayan yarasalar ile Eosen fosillerindeki yarasaların önayak parmaklarının göreceli orantıları kıyaslandığında önemli farklılıklar görülmemesi yarasa kanat morfolojisinin 50 milyon yıldan üzün bir süredir değişmediğini göstermektedir.[13] Uçuş sırasında kemikler hem bükülme hem de kayma gerilimine maruz kalır. Bükülme gerilimleri karasal memelilerden daha azdır ancak kayma gerilimi daha fazladır. Yarasaların kanat kemiklerinin kopma gerilim noktası kuşların kanat kemiklerinden biraz daha düşüktür.[14]
Diğer memelilerde olduğu gibi ve kuşların aksine önkolun ana bileşeni radiustur. Yarasaların hepsi bilekten çıkan beş uzamış parmağı bulunur. Başparmak ileri doğru bakar ve kanadın hücum kenarını desteklerken diğer parmaklar kanat zarındaki gerilimi desteklerler. İkinci ve üçüncü parmaklar kanat ucu boyunca uzanır ve aerodinamik sürükleme kuvvetine karşı kanadın ileri doğru çekilmesine yardımcı olurlar ve bu şekilde kanatlar teruzor kanadı kadar kalın olmak zorunda değildir. Dördüncü ve beşinci parmaklar bilekten firar kenarına doğru gider ve gergin zara doğru havanın itmesiyle oluşan bükülme kuvvetini iter.[15] Esnek eklemleri sayesinde yarasalar süzülerek uçan memelilerden daha iyi manevra yapabilir ve daha beceriklidirler.[16]

Yarasaların kanatları kuşların kanatlarından çok daha incedir ve daha çok kemik içerir ve böylece daha çok kaldırma ve daha az sürüklenme ile kuşlardan daha iyi manevra yapabilerek uçabilirler.[17] Kanat çırparken yukarıya doğru hareket sırasında kanatlarını vücutlarına doğru toplamaları uçuş sırasında %35 enerji korumalarını sağlar.[18] Kanat zarları hassastır ve kolaylıkla yırtılabilir[19] ama tekrar büyüyebilir ve küçük yırtıklar çabuk iyileşir.[19][20] Kanatların yüzeyinde küçük yumrular üzerinde dokunmaya duyarlı ve Merkel hücresi denen duyu organları vardır; bu hücreler insanların parmak ucunda da bulunur. Bu duyarlı alanlar yarasalarda farklıdır; her yumrunun merkezinde ince bir kıl bulunur ve böylece hava akışını algılayıp değişikliğine uyum sağlamaya izin verir. Asıl yararları uçmak için en etkin hızı tespit etmek ve perdövitesi engellemektir.[21] Böcekçil yarasalar bu taktil kılları uçarken avlarını yakalamak için karmaşık manevralar yaparken de kullanıyor olabilir.[16]
Kanat zarına patagium adı verilir. Kanat zarı kol ve parmak kemiklerinin arasında gerilmiştir ve gövdenin yanları boyunca arka ayaklara ve kuyruğa kadar iner. Bu deri zar bağ doku, elastik fibriller, sinirler, kaslar ve damarlardan oluşur. Kaslar uçuş sırasında zarı gergin tutar.[22] Kuyruğun patagiuma bağlı olmas seviyesi türler arasında farklılık gösterir; bazılarının tamamen serbest kuyrukları olduğu gibi bazıları tamamen kuyruksuzdur.[8] Bir kat epidermis ve dermis tabakası ile kıl kökleri ile ter bezlerine sahip yarasanın gövdesindeki deri kanat zarındaki deriden çok farklıdır. Patagium oldukça ince çifte epidermis katından ıluşur ve bu katların arasında kolajen ile elastik fibriller açısından zengin bir bağ doku bulunur. Parmakların arası dışında zarın üzerinde ne kıl kökü ne de ter bezi bulunur.[21][23] Yarasa embriyoları için apoptozis (hücre ölümü) yalnızca arka ayakları etkiler önayaklar ise daha sonra kanat zarına dönüşecek olan parmaklar arasındaki perdeleri korumaya devam eder.[24] Bükülmez kanatlarının bükülme ve burulma gerilimlerini omuzlara aktardığı kuşların aksine yarasaların esnek kanat zarları sadece gerilmeye dayanır. Uçabilmek için yarasa kanat zarının iskelet ile bağlandığı noktalara içeri doğru bir güç uygulamak zorundadır. Bu şekilde kanat uçlarında kanat yüzeyine dik gelen karşı kuvvet dengelenir. Bu adaptasyon uçuş sırasında kanatlarını kısmen katlayıp yukarı doğru kanat vuruş sırasında kanar açıklığını ve alanını azaltabilen ve süzülebilen kuşların aksine yarasalar kanat açıklıklarını azaltamazlar. Dolayısıyla da yarasalar kuşlar kadar uzun mesafe uçamazlar.[15]
Nektar ve polen ile beslenen yarasalar, sinek kuşlarına benzer şekilde uçarken havada asılı kalabilirler. Kanatların keskin hücüm kenarlarının yarattığı girdaplar kaldırma sağlar. Kanat eğriliklerini değiştirerek hayvan bu girdapları dengede tutabilir.[25]
Tüneme ve yürüyüş şekilleri

Uçmadıkları zaman yarasalar ayaklarından tutunarak başaşağı tüneme olarak bilinen pozisyonda dururlar.[26] Femurlar kalçalara, uçarken dışarı ve yukarı bükülebilecek şekilde bağlıdır. Arka ayak bileği eklemi kanatların firar kenarının aşağı doğru bükülmesine olanak sağlayacak şekilde esneyebilir. Bu ağaç dallarından aşağı sarkmak ya da tırmanmaktan başka harekete izin vermez.[15] Büyük yarasaların çoğu kafaları karınlarına doğru kıvrılmış şekilde tünerken küçük yarasaların çoğu boyunları sırtlarına doğru bükülmüş şekilde tüner. Bu farklılık her iki grubun boyun omurlarının yapısında açıkça görülür.[26] Yarasalar tünerken tendonlar ayaklarını kapalı olarak tutmalarına olanak sağlar. Tünekten kalkmak için ayakları açarken kas gücü gereklidir ama tutunmak için bu şekilde kas gücüne gerek kalmaz.[24]
Yerde iken yarasaların çoğu çok hantal bir şekilde yürür. Yalnızca Mystacina tuberculata ve Desmodus rotundus gibi birkaç tür yerde çevik hareket edebilir. Her iki tür de yavaş yavaş yürürken adımlarını birer birer atar ancak daha hızlı giderken katlanmış kanatlarını ileriye atılmak için kullanan D. rotundus iki ayağıyla birden sıçrayarak ilerler. D. rotundus türünün bu hareket şeklini muhtemelen yerde konaklarını izlemek için evrimleşmiş olması muhtemelken M. tuberculata türünün ise bölgelerinde rakip memeli türleri olmaması nedeniyle evrimleşmiş olması muhtemeldir.[27]
Dahili sistemler
Yarasalar etkili bir dolaşım sistemine sahiptir. Toplardamar çeper kaslarının ritmik kasılmasından ibaret oldukça güçlü bir toplardamar hareketini kullandıkları dikkati çeker. Memelilerin çoğunda toplardamarların çeperleri yalnızca pasif bir direnç gösterir ve oksijeni alınmış kan içlerinden akarken şekillerini korur ancak yarasalarda toplardamarların bu pompalama eylemiyle kanın kalbe geri akışını aktif olarak desteklediği görülmektedir.[28][29] Gövdeleri görece küçük ve hafif olduğu için yarasalar tünediklerinde kafalarına kan hücum etmesi riski taşımazlar.[30]
Yarasalar, büyük bir miktarda sürekli oksijen akımına ihtiyaç duyan dolayısıyla da çok enerji harcayan uçma eyleminin gerekliliklerini sağlamak için oldukça iyi uyum sağlamış bir solunum sistemine sahiptir. Yarasalarda orantısal olarak alveoler yüzey alanı ve akciğer kılcal damar kan hacmi diğer küçük dört ayaklı memelilerin çoğundan daha fazladır.[31] Uçuş sırasında solunum döngüsü kanat çırpma döngüsü ile bire bir bağıntı içindedir.[32] Memeli akciğerlerinin kısıtları yüzünden yarasalar yüksek rakım uçuşlarına dayanamazlar.[15]

Yarasaların uçma kaslarının çalışması için çok enerji ve verimli bir dolaşım sistemi gerekir. Uçuş için kullanılan kasların ihtiyaç duyduğu enerji, uçma dışında yollarla hareket eden memelilerin kaslarının ihtiyaç duyduğu enerjinin iki katıdır. Bu enerji tüketimine paralel olarak da uçan hayvanların kanlarındaki oksijen düzeyi karada hareket eden hayvanların iki katı kadardır. vücudun tamamına sağlanan oksijen miktarını kan ikmali kontrol ettiği için dolaşım sistemi de buna uygun olarak cevap vermelidir. Dolayısıyla aynı orantıda bir vücuda sahip karada yaşayan bir memelinin kalbine göre bir yarasanın kalbi üç kat daha fazla büyük olabilir ve daha fazla kan pompalar.[34] Kalp debisi doğrudan nabız hızı ve atım hacmine bağlıdır;[35] aktif bir küçük yarasanın nabzı dakikada 1000 kere atabilir.[36]
Oldukça ince zar dokusuyla, bir yarasanın kanadı organizmanın toplam gaz değişimi verimliliğine önemli ölçüde katkıda bulunur.[23] Uçmanın yüksek enerji ihtiyacı nedeniyle yarasanın vücudu bu ihtiyacı kanatların patagiumundan gaz değişimi ile karşılar. Yarasanın kanatları açık olduğunda yüzey alanının hacme olan oranında artış olur. Kanatların yüzey alanı toplam vücut yüzey alanının %85'i kadardır dolayısıyla da yararlı miktarda gaz değişimi olasılığını gösterir.[23] Kanat zarında deri altı damarlar yüzeye çok yakındır ve oksijen ile karbondioksitin difüzyonuna olanak sağlarlar.[37]
Yarasaların sindirim sistemi türüne ve beslenme tarzına göre farklı adaptasyonlara sahiptir. Diğer uçan hayvanlarda olduğu gibi enerji gereksinimi ile başa çıkabilmek için besin hızlıca ve verimli bir şekilde işlenir. Böcekçil yarasalar, yedikleri böcekleri daha iyi sindirebilmek için bazı sindirim enzimlerine sahiptir; örneğin böceklerin büyük bir kısmını oluşturan kitini çözebilmek için kitinaz gibi.[38] Vampir yarasalar, muhtemelen kan ile beslendikleri için sindirim sistemlerinde malt şekerini çözen maltaz enzimine sahip olmayan tek omurgalılardır. Nektarcıl ve meyvecil yarasalar gıdalarındaki yüksek şeker miktarı nedeniyle böcekçillerden daha fazla maltaz ve sükraz enzimine sahiptirler.[39]
Yarasaların böbrekleri de beslenmelerine göre adaptasyona uğramıştır. Etçil ve vampir yarasalar yüksek oranda protein tükettikleri için yoğuşuk idrar çıkarabilirler; böbreklerinin ince bir korteksi ve uzun böbrek kabarcıkları olur. Meyvecil yarasalarda bu özellik yoktur ve böbrekleri düşük elektrolitli gıdalarla beslenmeleri nedeniyle elektrolit tutucu olarak uyum sağlamışlardır.; buna uygun olarak böbreklerinin korteksi kalındır ve çok kısa oknik kabarcıkları bulunur.[39] Yarasalar uçmaya bağlı olarak daha yüksek metabolik hızlara sahiptir ve dolayısıyla da solunum ile su kaybı da daha yüksektir. Oldukça damarlı zardan oluşan geniş kanatları yüzey alanını artırır ve deriden buharlaşma yolu ile su kaybına neden olur.[33] Su kanlarında ve termoregülasyon sistemlerinde iyonik dengeyi sağlamaya ve atık ile toksinlerin idrar yolu ile vücuttan atılmasına yardımcı olur. Eğer yeteri kadar sıvı almazlarsa kan üre azotu zehirlenmesine uğrayabilirler.[40]
Dişi yarasalarda rahim yapısı da türlere göre değişiklik gösterir, bazılarının iki rahim boynuzu bulunurken bazılarında tek bir anahat odacığı vardır.[41]
Duyular
Ekolokasyon

Küçük yarasalar ile büyük yarasaların bazıları yankı üretmek üzere ultrasonik ses yayarlar. Bu yankıların ses şiddeti subglottik basınca bağlıdır. Darbe frekansının oryantasyonunu kontrol eden yarasaların krikotiroid kasının işlevi önemlidir. Bu kas gırtlağın içindedir ve ses çıkarmaya yardımcı olabilen tek tensor kastır.[42] Giden darbelerle dönen yankıları kıyaslayarak beyin ve işitme sisnir sitemi yarasanın çevresinin detaylı imajlarını üretebilir. Bu şekilde yarasalar karanlıkta avlarını algılayıp, yerini tespit eder ve sınıflandırabilirler. Yarasa çağrıları uçan hayvan seslerinin en yükseklerinden biridir ve şiddeti 60 ila 140 desibel arasında değişir.[43][44] Küçük yarasalar ultrason yaratmak için gırtlaklarını kullanır ve bunu ağızlarından, bazen de burunlarından yayarlar. Burundan yayma nalburunlu yarasalarda (Rhinolophus spp.) daha belirgindir. Küçük yarasa çağrılarının frekansı 14.000 Hz'den 100.000 Hz.'in üzerine kadar çıkabilir ki insan duyma sınırları 20 ila 20.000 Hz. arasındadır.[45] Çeşitli yarasa grupları burun deliklerinin çevresinde ve üzerinde burun yaprağı denen ve ses iletiminde rol oynayan etsi uzantılar evrimleştirmiştir.[24]

Düşük görev çevrimli ekolokasyonda yarasalar çağrılarını ve yankıları zamana göre ayırabilirler. Kısa çağrılarını yankılar dönmeden bitirecek şekilde zamanlamalıdırlar. Yarasalar çağrı yaparken orta kulak kaslarını kasarak kendilerini sağır etmezler. Çağrı ile yankı arasındaki zaman farkı bu kasları gevşetip gelen yankıyı duymalarına olanak verir.[46] Dönen yankıların gecikme süreleri yarasaya avlarının uzaklığını kestirmeye yardımcı olur.[45]
Yüksek görev çevrimli ekolokasyonda yarasalar sürekli çağrı yapar ve darbe ile yankı frekansını ayırırlar. Bu yarasaların kulakları belirli bir frekans aralığına ayarlıdır. Yaptıkları çağrı bu frekans aralığının dışındadır ve bu şekilde kendilerini sağır etmezler. Daha sonra uçarken hareket ettikleri için oluşan Doppler etkisinden yararlanarak gelen yankıları duyabildikleri frekans aralığında alırlar. Dönen yankıların Doppler etkisi avlarının hareketş ve yeri hakkında bilgi taşır. Bu yarasalar uçuş hızlarındaki değişikliğin Doppler etkisi üzerindeki değişikliği ile başa çıkmak zorundadırlar. Yaydıkları ses darbe frekansını uçuş hızlarına göre değiştirmeye uyum sağlamışlardır ve böylece yankılar hep optimal duyma aralığında geri gelir.[47]
Avlarının yerini ekolokasyonla belirlemenin yanı sıra yarasa kulakları güve kanatlarının çırpışmasına, timballi böceklerin çıkardığı seslere ve kırkayaklar ile kulağakaçanlar gibi yerde dolaşan böceklerin hareketlerinin çıkardığı seslere duyarlıdır. Yarasaların kulaklarının iç yüzeyindeki yükseltilerin karmaşık geometrisi ekolokasyon sinyallerini odaklamaya ve avların çıkardığı diğer sesleri pasif olarak dinlemeye yardımcı olur. Bu yükseltilere Fresnel lensin akustik eşleniği gözüyle bakılabilir ve birbirleriyle ilişkisi bulunmayan ay-ay, Afrika küçük lemuru, iri kulaklı tilki, fare lemuru ve başka hayvanlarda da görülür.[48][49][50] Yarasalar yankıların dış kulaklarında bir deri çıkıntısı olan tragustan yansımasıyla oluşan girişimleri kullanarak avlarının yerden yüksekliğini kestirebilirler.[45]

Tekrar tekrar tarama yaparak yarasalar içinde hareket ettikleri çevre ile avlarının doğru bir imajını zihinlerinde oluşturabilirler.[53] Kaplan güveleri gibi bazı güve türleri yarasalara kimyasal olarak korundukları ve dolayısıyla da lezzetsiz oldukları uyarısını veren aposematik ultrason sinyalleri yaratarak yarasaların bu özelliklerinden yararlanmışlardır.[51][52] Kaplan güveleri de dahil olmak üzere bazı güve türleri yarasaların ekolokasyonunu bozmak üzere sinyallar üretebilir. Birçok güve türünün gelen yarasa sinyalinin güvenin uçuş kaslarını düzensizce çalıştırmaya neden olan ve güveyi gelişigüzel kaçış manevralarına sürükleyen timpanal organı adı verilen bir işitme organları vardır.[54][55][56]
Görme
Çoğu küçük yarasa türünün gözleri küçük ve pek gelişmemiştir dolayısıyla görme keskinliği zayıftır ama hiçbir tür kör değildir.[57] Küçük yarasaların çoğu mezofik görme sahibidir yani düşük düzeyde ışığı algılayabilirler halbuki diğer memeliler renkleri görmelerini sağlayan fotopik görme sahibidir. Ekolokasyon yalnızca kısa mesafelerde etkili olduğu için küçük yarasalar görme duyularını tünekleri ve avlanma yerleri arasında gitmek için kullanırlar. Bazı türler morötesini algılayabilir. Bazı küçük yarasaların gövdeleri farklı renklere sahip olduğu için belki renkleri ed ayırt edebilmektedirler.[11][58][59][60]
Büyük yarasalarınsa görme yetenekleri en az insanlar kadar iyi hatta belki de daha iyidir. Görüşleri hem gece hem de gündüz görüşüne uyum sağlamıştır ve biraz renkli de görebilirler.[60]
Manyetoalgılama
Küçük yarasalar manyetoalgılama kullanılırlar; kuşlar gibi Dünya'nın manyetik alanına büyük bir hassasiyetleri vardır. Küçük yarasalar kutup bazlı pusula kullanırlar yani kuzey ile güneyi ayırt edebilirler halbuki kuşlar manyetik alanın gücünü kullanarak uzun mesafe yolculukta kullanılmak üzere enlemleri ayırt edebilirler. Nasıl işlediği bilinmemekle birlikte manyetit partiküllerinin rolü olabilir[61][62]
Termoregülasyon

Yarasaların çoğu homeotermiktir yani kararlı bir vücut sıcaklığına sahiptir, bunlara istisna heterotermik olan yani vücut sıcaklıkları değişiklik gösteren vesper yarasaları (Vespertilionidae), nalburunlu yarasalar (Rhinolophidae), serbest kuyruklu yarasalar (Molossidae) ve bükük kanatlı yarasalardır (Miniopteridae).[63] Diğer memelilere kıyasla yarasaların ısıl iletkenliği yüksektir. Kanatlar kan damarları ile kaplıdır ve açıldıklarında ısı kaybederler. Dinlenirken kanatlarını vücutları etrafında sararak ılık havayı hapsederler. Küçük yarasaların metabolizma hızları büyük yarasalardan daha yüksektir ve homeotermiyi sağlamak için daha fazla besin almak zorundadırlar.[64]
Koyu renkli kanatları güneş radyasyonunu soğurduğu için güneş altında aşırı ısınmayı engellemek için gündüzleri uçmaktan kaçınırlar. Eğer ortam sıcaklığı çok yüksekse yarasalar ısıyı dağıtamayabilirler[65] aşırı koşullar altında kendilerini serinletmek için salyalarını kullanırlar.[15] Büyük yarasalar arasında uçan tilki Pteropus hypomelanus günün en sıcak kısmında tünediği yerde salyasını ve kanatlarını kullanarak kendini serinletir.[66] Küçük yarasalar arasında Myotis yumanensis, Tadarida brasiliensis ve Antrozous pallidus türleri soluyarak, salya çıkararak ve buharlaşma ile serinlemeyi sağlamak için kürklerini yalayarak 45 °C sıcaklığa kadar dayanabilirler; bu şekilde metabolik ısı üretimlerinin iki katını kaybedebilirler.[67]
Yarasaların ayrıca kanatlarının kenarları boyunca dolanan damar ağlarının atardamar kısmında bir büzgen valf sistemi bulunur. Bunlar tam olarak açık olduğunda oksijenli kanın kanat zarı üzerinde kılcal damar ağı boyunca dolaşmasına izin verirken kapalı olduklarında da şant akımı kanat kılcal damarlarına uğramadan toplardamarlara gider. Bu şayede yarasalar kanat zarından ne kadar ısı transferi yapılacağını kontrol eder ve uçuş sırasında ısıyı dışarı bırakırlar. Başka memelilerin birçoğu büyük kulaklarındaki kılcal damar ağını aynı iş için kullanır.[68]
Uyuşukluk

Vücut sıcaklığının ve metabolizmanın azaldığı fiziksel aktivitenin azalma durumu olan uyuşukluk aktif iken çok büyük miktarda enerji harcayan, güvenilmez besin kaynaklarına bağlı ve yağ depolamak için sınırlı imkanları olan küçük yarasalar için özellikle yararlıdır. Bu durumda iken vücut sıcaklıklarını 6-30 °C'ye kadar düşürebilir ve enerji harcamalarını %50 ila %99 arasında azaltabilirler. Küçük yarasaların yaklaşık %97'si uyuşukluğa girer.[69] Tropik yarasalar avlanmak ile geçirdikleri zamanı azaltmak ve böylece de kendilerinin av olması şansını azaltmak için uyuşukluğa girebilirler.[70] Büyük yarasaların genel olarak homeotermik olduğuna inanılır ancak bunların içinde yaklaşık 50 g. ağırlıkları ile küçük boyutta olanların uyuşukluğa girdiği bilinmektedir: Syconycteris australis, Macroglossus minimus ve Nyctimene robinsoni. Büyük yarasalar için uyuşukluk durumu yazları kışları olduğundan daha uzun sürer.[71]
Kış uykusu sırasında yarasalar bir uyuşukluk durumuna girer ve bu dönem boyunca vücut sıcaklıklarını %99,6 kadar düşürürler; hatta uyanma dönemleri sırasında vücut sıcaklıkları normale döndüğünde bile bazen "heterotermik uyanma" olarak bilinen yüzeysel bir uyuşukluk durumuna girerler.[72] Bazı yarasalar yaz aylarında serin kalmak için yüksek sıcaklıklar sırasında uyur duruma geçerler.[73]
Uzun göçler sırasında heterotermik yarasalar geceleri uçup gündüzleri tünerken uyuşukluğa girer. Gündüz uçup gece beslenen göçmen kuşların aksine gececil yarasalar yol yapmak ve beslenmek arasında çelişkiye düşerler. Tasarruf ettikleri enerji beslenme ihtiyaçlarını azaltmasının yanı sıra göçün süresini de azaltır ve bilmedikleri yerlerde daha fazla zaman geçirmeyip avlanmalarını da azaltır. Bazı türlerde gebe bireyler uyuşukluğa girmeyebilir.[74][75]
Boyutları
En küçük yarasa 29 ila 34 mm uzunluğunda ve 15 cm kanat açıklığına sahip olan, 2 ila 2,6 g ağırlığında olan yabanarısı yarasasıdır (Craseonycteris thonglongyai).[76][77] Ayrıca cüce sivri farenin yanında en küçük yaşayan memeli türü olduğu da iddia edilir.[78] En büyük yarasalar Pteropus büyük yarasaların birkaç türü ile 1,6 kg ağırlığında ve 1,7 m kanat açıklığına sahip olan Acerodon jubatus türü yarasadır.[79] Ekolokasyon için büyük yarasalar düşük frekansları, küçük yarasalar da yüksek frekansları kullanmaya meyillidir; küçük avların yerini tespit etmek için yüksek frekanslı ekoloksayon daha etkilidir. Tespit etmeleri mümkün olmadığı için küçük avlar büyük yarasalar tarafından yenmez.[80] Bir yarasa türünün sağladığı uyumlar hangi avları avlayabileceğini doğrudan etkilemektedir.[81]
Ekoloji

Uçma yetenekleri sayesinde yarasalar en yaygın dağılmış memeli gruplarından biri olmuştur.[82] Arktik'in yüksek enlemleri, Antarktika ve izole birkaç okyanus adasının dışında yarasalar Dünya'nın hemen hemen her habitatında yaşarlar.[83] Tropik alanlar ılıman alanlardan daha fazla türe sahiptir.[24] Farklı türler farklı mevsimlerde sahillerden dağlara ve çöllere kadar farklı habitatlar seçerler ama tüneyebilecekleri yerlere ihtiyaç duyarlar. Yarasa tünekleri kovuklarda, yarıklarda, bitki örtüsü içinde ve hatta insan yapısı yapılarda rastlanabilir; bazı yarasa türleri yapraklar ile kendilerine "çadır" bile kurmaktadır.[84] Büyük yarasalar genellikle ağaçlara tünerler.[85] Küçük yarasaların çoğu gececilken[86] büyük yarasal tipik olarak gündüzleri ya da alacakaranlıkta aktiftirler.[87][88]
Ilıman alanlarda bazı küçük yarasalar kış uykusu barınaklarına doğru yüzlerce kilometre göç ederler.[89] Bazılar soğuk havada uyuşukluk durumuna geçer ve ılık hava böceklerin aktif olmasına izin verdiği sürece uyuşukluktan çıkıp avlanırlar.[89] Bazıları da kış için mağaralara çekilip altı ay kadar kış uykusuna yatar.[89] Ekolokasyonlarını etkilediği ve avlanamadıkları için küçük yarasalar nadiren yağmur altında uçar.[90]
Besin ve beslenme
Farklı yarasa türleri aralarında böcekler, nektar, polen, meyve ve hatta omurgalılar gibi farklı besinlerle beslenirler.[24] Büyük yarasalar çoğunlukla meyve, nektar ve polenle beslenir.[87] Küçük boyutları, yüksek metabolizmaları ve uçuş sırasında hızlı enerji tüketimi nedenleriyle yarasalar boyutlarına göre çok miktarda besin tüketmek zorundadır. Böcekçil yarasalar vücut ağırlıklarının %120'sinden fazlasını yiyebilirken meyvecil yarasalar ağırlıklarının iki katından fazlasını yiyebilir.[24] Besin peşinde geceleri önemli mesafeler katedebilirler, istisnai olarak Euderma maculatum 38,5 km. gidebilmektedir.[91] Yarasalar çeşitli avlanma stratejileri kullanırlar.[80] Yarasalar ihtiyaç duydukları suyun çoğunu yedikleri besinlerden alırlar; çok sayıda türde göller ve ırmakların üzerinden uçarken dillerini suya batırarak da su içer.[24]
Chiroptera takımı bir bütün olarak C vitamini sentezleme yeteneğini kaybetme süreci içindedir.[92] Başlıca böcekçil ve meyvecil yarası familyalarının da aralarında bulunduğu altı ana familyadan 34 yarasa türü üzerinde yapılan test sonucunda hepsinin C vitamini sentezleme yeteneğini kaybettiği görülmüştür ve bu kayıp tek bit mutasyonla ortak bir atadan gelmiş olabilir.[93][b] En azından iki yarasa türü meyvecil Rousettus leschenaultii ve böcekçil Hipposideros armiger C vitamini üretme yeteneğini korumuştur.[94]
Böcekler
Küçük yarasaların çoğu, özellikle de ılıman alanlarda olanlar böceklerle avlanırlar.[24] Böcekçil yarasalar çok çeşitli böceklerle beslenebilirler,[24] bunların arasında sinekler, sivrisinekler, kın kanatlılar, güveler, çekirgeler, cırcır böcekleri, termitler, arılar, yaban arıları, mayıs sinekleri ve şayak sinekleri sayılabilir.[8][95] ABD'nin Teksas eyaletinin ortasında çok sayıda Tadarida brasiliensis türü yarasa yerden yüzlerce metre yüksekte birlikte uçarak göç eden güvelerle beslenirler.[96] Böcekleri uçarken yakalayan Myotis lucifugus gibi türler havada iken böcekleri ağzı ile yakalayıp havada iken yiyebilir ya da kuyruk veya kanat zarları ile böceği yakalayıp ağızlarına götürebilir.[97][24] Yarasa yakaladığı böceği tüneğine götürüp orada da yiyebilir.[98] Plecotus auritus türü ve nalburunlu yarasa türlerinin çoğu gibi yavaş hareket eden yarasa türleri böcekleri bitkilerin üzerinde iken avlayabilir.[8] Yüksek enlemlerde yaşayan böcekçil yarasalar tropik yarasalardan daha yüksek enerji değerine sahip avlarla beslenmek zorundadır.[99]
Meyveler ve nektar

Meyve yiyen yarasalar iki ana alt takımda da bulunur. Yarasalar ağaçtan dişleriyle çekip kopardıkları olgun meyveleri tercih ederler. Meyveyi yemek için tüneklerine geri uçarlar. Meyvenin suyunu emer ve tohumları ile posasını yere tükürürler. Bu meyve ağaçlarının tohumlarının yayılmasına yardımcı olur. Bu tohumlar yarasaların bıraktıkları yerlerde filizlenip kök salabilir. Birçok bitki türü tohum dağılımı için yarasalara bağlıdırlar.[100][24] Artibeus jamaicensis türü yarasaların 3 ila 14 g ve hatta 59 g ağırlığında meyveleri taşıdıkları kaydedilmiştir.[101]
Nektar yiyen yarasaların kendilerine özgü adaptasyonlar geçirmişlerdir. Bu yarasaların uzun burunları, belirli çiçekler ve bitkilerden beslenmelerine yardımcı olan ince kıllarla kaplı, uzun ve uzanabilen dilleri vardır.[24][102] Anoura fistulata türü yarasa memeliler arasında vücut boyutlarına oranla en uzun dile sahip hayvandır. Polenleşme ve beslenme için bu yararlı olmaktadır. Uzun ve dar dilleri bazı uzun kadeh şekilli çiçeklerin derinlerine ulaşmalarına yardımcı olmaktadır. Dil geri çekildiğinde göğüs kafesinin içinde kıvrılarak toplanır.[102] Bu özelliklerinden ötürü nektarla beslenen yarasalar besin kaynaklarının azlığı durumunda kolaylıkla başka besin kaynaklarına dönemezler ve bu da onları diğer yarasa türlerinden daha fazla soy tükenme tehlikesine sokar.[103][104] Nektarla beslenme ayrıca çeşitli bitkilere yardımcı da olmaktadır çünkü beslenirken kürklerine bulaşan polenler nedeniyle polen taşıyıcı olarak da yardımcı olurlar. Yaklaşık 500 çiçek türü yarasaların polenlemeye yardımcı olmasına muhtaçtırlar ve bu nedenle geceleri çiçek açarlar.[24] Birçok yağmur ormanı yarasaların polen taşıyıcılığına ihtiyacı vardır.[105]
Omurgalılar

Bazı yarasalar balıklar, kurbağalar, kertenkeleler, kuşlar ve memeliler gibi diğer omurgalıları avlarlar.[8][24] Örneğin Trachops cirrhosus kurbağaları yakalama konusunda çok beceriklidir. Bu yarasalar, çiftleşme çağrılarını takip ederek büyük kurbağa gruplarının yerini tespit eder ve keskin köpek dişleriyle su yüzünden kurbağaları toplarlar.[107] Nyctalus lasiopterus uçarken kuşları yakalayabilir.[106] Noctilio leporinus gibi bazı türler balık avlar. Su yüzündeki küçük dalgalanmaları ekolokasyon ile tespit edebilir ve sonra birdenbire inerek arka ayaklarında özel olarak büyümüş pençeleriyle balığı yakalar ve tünekleri götürerek yerler.[108] En azından iki yarasa türünün diğer yarasalarla beslendiği bilinmektedir: Vampyrum spectrum ve Macroderma gigas.[89]
Kan

Özellikle Diaemus youngi ve kıllı bacaklı vampir yarasa (Diphylla ecaudata) vampir yarasaları olmak üzere birkaç tür yalnızca hayvan kanı ile beslenir. Desmodus rotundus genellikle sığır gibi büyük memelilerden beslenirken yukarıda belirtilen diğer iki yarasa kuşların kanıyla beslenir.[109] Vampir yarasalar uyuyan avları hedeflerler ve derin soluk almayı algılarlar.[24] Burunlarındaki ısı algılayıcıları derinin yüzeyine yakın kan damarlarını tespit etmelerine yardım eder.[110] Dişleriyle hayvanın derisini deler ve küçük bir kısmını kaldırırlar[111] ve bu iş için uyum geçirmiş yanal kanalcıklara sahip dilleriyle çıkan kanı yalarlar.[112] Salyalarındaki antikoagülan kanın pıhtılaşmasını engeller.[111]
Avcılar, parazitler ve hastalıklar
Yarasalar baykuşlar, atmacalar ve doğanlar gibi yırtıcı kuşlar ve tüneklerinde de kediler gibi tırmanabilen hayvanlar tarafından avlanırlar.[113] Yirmi kadar Yeni Dünya yılan türünün yarasaları yakaladıkları bilinmektedir; bu yılanlar mağaralar gibi yarasaların sığındıkları yerlerin girişlerinde bekleyerek uçan yarasaları yakalarlar.[114] J. Rydell ve J. R. Speakman yarasaların erken ve orta Eosen döneminde avcılardan kaçınabilmek için gececil olarak evrimleştiklerini öne sürerler.[113] Bazı zoologlar şu ana kadar kanıtların müphem olduğunu düşünmektedir.[115]

Yarasaların taşıdığı ektoparazitler arasında pireler ve akarlar dışında yarasalara özgü yarasa böcekleri ve yarasa sinekleri (Nycteribiidae ve Streblidae) sayılabilir.[116][117] Yarasalar bitlere konaklık etmeyen suda yaşamayan birkaç memeli takımı arasında yer alır. Bu muhtemelen aynı nişi dolduran özellişmiş asalaklarla olan rekabet yüzündendir.[117]
Beyaz burun sendromu ABD'nin doğusunda ve Kanada'da milyonlarca yarasanın ölümüne neden olan bir durumdur.[118] Hastalağın adı yakalanan yarasaların burunlarında, kulaklarında ve kanatlarında büyüyen beyaz Pseudogymnoascus destructans mantarından gelir. Manatar muhtemelen yarasadan yarasaya geçmekte ve hastalığa neden olmaktadır.[119] Mantar ilk olarak 2006'da New York Eyaleti'nin orta kısımlarında keşfedildi ve kısa sürede Florida'nın kuzeyinde tüm doğu sahillerine yayıldı; ölüm oranları en çok etkilenen mağaralarda %90 ila %100 arasında olarak tespit edildi.[120] 2006 yılından beri New England ve Orta Atlantik eyaletlerinde bazı türlerin tamamı yok oldu ve bazı türlerin sayıları da yüzlerce bin ve hatta milyonlarca yarasadan yalnızca birkaç yüz ve daha az yarasaya indi.[121] Kanada'nın Nova Scotia, Quebec, Ontario ve New Brunswick eyaletlerinde de benzer ölümler görüldü ve Kanada hükûmeti tüm topraklarında kalan yarasa popülasyonunu korumak için hazırlıklar yapmaya başladı.[122] Bilimsel kanıtlar mantarın yarasalara bulaşmak için daha uzun bir döneme sahip olduğu uzun kışlar daha büyük ölüm sayısıyla sonuçlanmaktadır.[123][124][125] 2014 yılında hastalık Mississippi Nehri'ni aştı[126] ve 2017'de Teksas'ta yarasalar üzerinde görüldü.[127] 2016 yılının Mart ayında ABD'nin doğusunda Washington eyaletinde bir yarasa üzerinde bulunduğu tespit edildi.[128]
Yarasalar aralarında birçok yarasa popülasyonunda endemik olarak bulunan kuduz,[129][130][131] hem doğrudan hem de guano yoluyla histoplazmozis,[132] Nipah ve Hendra virüsleri,[133][134] ve muhtemelen ebola virüsü[135][136] gibi çok sayıda zoonoz patojenin doğal konağıdır.[137] Yüksek hareketlilikleri, yaygın dağılımları, uzun yaşam süreleri, türlerin önemli ölçüde simpatrisi (yaşam alanı örtüşmesi) ve sosyal davranışları yarasaları hastalıklar için elverişli konak ve vektör hâline getirmektedir. Kemiricilerle kıyaslandığında yarasalar tür başına daha çok zoonoz virüs taşırlar ve her virüs daha çok tür ile paylaşılır.[138] Taşıdıkları patojenlerin çoğuna yüksek oranda dirençli oldukları görünmektedir ve bu da bağışıklık sistemlerinin adaptasyon düzeyini göstermektedir.[138][139][140] Çiftlik ve ev hayvanları ile etkileşimleri zoonotik hastalık geçiş riskini oluşturur.[130] Koronavirüslerin doğal konakları olduklarından ve Yünnan'da tek bir mağaradan gelen yarasaların Çin'de şiddetli akut solunum yolu sendromunun (SARS) çıkışının nedeni olarak gösterilmektedir.[132][141][142]
Sosyal davranış
Sosyal yapı
Bazı yarasalar yalnız yaşarken diğerleri bir milyon bireyden fazla koloniler hâlinde yaşarlar.[89] Büyük koloniler hâlinde yaşamak bireyler için avlanılma riskini azaltır.[8] Ilıman yarasa türleri kış uykusuna yattıkları yerlere sonbahar yaklaştıkça sürü olarak gider. Bu şekilde genç yarasalar kış uykusu geçirilen yerler gösterilir ve erişkinler de diğer gruplardaki yarasalarla çiftleşebilir.[24]
Çeşitli türler fisyon-füzyon topluluk yapısına sahiptir; çok sayıda yarasa tünemek için aynı yere toplanır ve avlanmak için küçük gruplara ayrılır. Bu topluluklar içinde yarasalar uzun süreli ilişkiler sürdürebilmektedir.[143] Bu ilişkilerin bazıları anasoylu dişiler ile onlara bağımlı yavrularından oluşur.[144] Desmodus rotundus gibi bazı türlerde ortak beslenme ve ortak temizlenme gibi sosyal bağları güçlendiren davranışlar görülür.[145][146]
İletişim
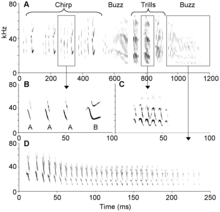
Yarasalar sesini en çok kullanan memeliler arasındadır ve eş bulmak, tünek arkadaşı bulmak ve kaynakları savunmak için çağrıda bulunur. Bu çağrılar genellikle düşük frekanslıdır ve uzun mesafeler katedebilir.[8][24] Tadarida brasiliensis türü yarasa kuşlar gibi şakıyabilen birkaç türden biridir. Erkekler dişileri cezbetmek için şakır. Şakımaları üç bölümden oluşur: Cıvıltı, titrek şakıma ve vızıltı. Yarasa şakımaları oldukça stereotipiktir ancak bireyler arasında hece sayısı, cümle sırası ve cümle tekrarı gibi farklılıklar içerir.[147] Phyllostomus hastatus türü yarasaların dişileri grubu bir arada tutmak için tünekdaşları arasında yüksek ve genişbantlı çağrılar yapar. Çağrılar tünek grupları arasında farklılık gösterir ve sesli öğrenme yoluyla ortaya çıkabilir.[148]
Esaret altındaki Mısır meyve yarasaları üzerine yapılan bir çalışmada kontrollü çağrıların %70'i araştırmacılar tarafından hangi yarasanın yaptığı anlaşılmaktadır ve çağrıların %60'ı dört kategoride değerlendirilebilir: besin üzerine tartışma, uyuma grubu içindeki konumları üzerine tartışma, çiftleşme girişimlerine karşı protesto ve birbirlerine yakın tünediklerinde tartışma. Hayvanlar, özellikle karşı cinsiyetten olmak üzere, farklı bireylerle iletişime girdiklerinde biraz farklı sesler çıkarmışlardır.[149] Yüksek oranda eşeysel dimorfizm gösteren Hypsignathus monstrosus türü yarasalarda ekekler dişileri çekmek için kalın, rezonanslı monoton çağrılar yapar. Uçan yarasalar trafik kontrolü için sesli sinyaller verir. Noctilio leporinus türü yarasa birbirleri ile çarpışacak bir yol üstünde uçuyorlarsa kaz gibi ses çıkarırlar.[24]
Yarasalar başka yollarla da iletişim kurarlar. Sturnira lilium türünün erkeklerinde üreme mevsiminde baharlı bir koku üreten omuz bezleri bulunur. Diğer çoğu tür gibi salgıları tutup dağıtmak için özelleşmiş kılları vardır. Böyle özelleşmiş kıllar bazı Eski Dünya büyük yarasalarının boyunlarında göze çarpan bir gerdanlık oluşturur. Saccopteryx bilineata türü yarasaların erkekleri kanatlarında bulunan keselerde salya ve idrar gibi vücut salgılarını birleştirilerek tünek yerlerine bırakılan parfüm gibi bir sıvı üretirler. Tüneklerin işaretlenmesi işlemine "tuzlama" denir ve aynı anda şakıdıkları da görülür.[24]
Üreme ve yaşam öyküsü

Stratejiler
Yarasa türlerinin çoğu polijindir yani erkekler birçok dişi ile çiftleşir. Pipistrellus ve Nyctalus cinsi yarasalar ile Desmodontinae alt familyasında sınıflandırılan yarasalar tünek yerleri gibi dişileri çekebilecek kaynaklara sahip çıkarak ve savunarak buralara gelen dişilerle çiftleşebilirler. Böyle yerlere sahip çıkamayan erkek yarasalar kenarda kalır ve üreme başarıları daha düşüktür.[8][150] Tadarida brasiliensis ve Myotis lucifugus gibi bazı türlerde ise hem erkek hem de dişi yarasa birçok eşle çiftleşir.[151][152] Bu yarasalarda dişilerin bazı erkeklere temayül ettiği görülmektedir.[8] Lavia frons ve Vampyrum spectrum birkaç türde ise erişkin erkek ve dişi yarasalar monogam çiftler oluşturur.[8][24] Erkeklerin bir araya gelip nümayiş yolu ile seçilen dişiler için rekabet ettileri "Lek çiftleşme" yarasalarda nadir rastlanır[153] ve Hypsignathus monstrosus türü yarasalarda görülür.[154]
Çiftleşme
Ilıman iklimlerde yaşayan yarasalar yaz sonu sonbahar başı arasında çiftleşirler.[155] Tropikal iklimde yaşayan yarasalar yağmurların kesildiği sezonda çiftleşebilirler.[156] Birleşmeden sonra erkek yarasa diğer erkeklerin spermlerini engellemek ve babalığını garanti etmek için arkada bir çiftleşme tıkacı bırakabilir.[157] Kış uykusuna yatan türlerde erkeklerin uyuşukluk içinde olan dişilerle çiftleştiği bilinmektedir.[8] Dişi yarasalar, doğum zamanını kontrol etme ve yavruların maksimum besin olduğunda doğmasını sağlamak için farklı stratejilere sahiptir. Bazı türlerin dişileri çiftleşmeden birkaç ay sonraya katar spermleri üreme organında saklayarak geçikmiş döllenme stratejisini kullanır. Çiftleşme sonbaharda olur ancak döllenme sonraki ilkbahara kadar gerçekleşmez. Bazı türler çiftleşme sonrası döllenmenin gerçekleştiği yumurtayı dış koşullar uygun olana kadar serbest hâlde üreme organlarında saklayarak embriyonik diyapoza girer ve döllenmiş yumurta daha sonra rahme tutunur.[155] Bir başka stratejide ise hem döllenme hem de rahme tutunma gerçekleşir ancak iyi koşullar sağlanana kadar fetusün gelişmesi geciktirilir. GEcikmiş gelişme sırasında anne döllenmiş yumurtayı besleyerek canlı tutar. Bu süreç gelişmiş gaz değişimi sistemi sayesinde uzun bir süre devam edebilir.[158]
Yaşam döngüsü

Ilıman bölgelerde yaşayan yarasalar genellikle Kuzey yarımkürede Mayıs ya da Haziran, Güney yarımkürede de Kasım ya da Aralık aylarında doğarlar. Tropikal bölgelerde doğum yağmur mevsiminin başında olur.[159] Türlerin çoğunda anne tek yavru taşır ve doğurur.[160] Doğum sırasında yavru yarasa annenin ağırlığının %40'ı kadar ağırlığa sahip olabilir[8] ve dişinin pelvis kemerinin iki yarısı esnek bir bağ doku ile bağlı olduğu için genişleyebilir.[89] Dişiler doğumu kolaylaştımak için yerçekimini kullanarak genellikle kafaları yukarıda ya da yatay konumda doğururlar. Yavru muhtemelen kanatların dolaşmasını engellemek için arkası önce konumda dışarı çıkar ve dişi yavruyu kanat ve kuyruk zarlarının içine alır. Türlerin çoğunda dişiler doğum kolonilerinde toplanarak bir arada doğururlar ve birbirlerine yardımcı olabilirler.[161][155][89]
Yavru yarasaların bakımının çoğunu anne üstlenir. Monogam türlerde baba da bunda rol alır. Bazı türlerde dişi başka yavruları da emzirebilir. Bu şekilde dişilerin üremek için doğdukları koloniye dönen türlerde koloni boyutları artış gösterir.[8] Genç bir yarasanın uçabilmesi için erişkin gövdesine ve ön ayak uzunluğuna erişmesi gerekir. Myotis lucifugus türünde bu doğumdan sonra on sekiz gün sonra gerçekleşir. Türlerin çoğunda yavruların sütten kesilmesi seksen günden önce olur. Desmodus rotundus yavrularını bu süreden sonra da emzirmeye devam eder ve gençleri diğer türlere göre daha sonra kendi başlarına bakabilecek hâle gelirler. Bunun nedeni muhtemelen kan temelli beslenmeleri ve her gece kan emmenin zorluğudur.[24]
Ortalama ömür
Yarasaların maksimum ömür uzunluğu benzer boyutlardaki diğer memelilerin üç buçuk katıdır. Altı türün doğal yaşam alanlarında 30 yıldan fazla yaşadığı kaydedilmiştir: Plecotus auritus, Myotis lucifugus, Myotis brandti, Myotis blythii, Rhinolophus ferrumequinum ve Pteropus giganteus.[162] Yaşam hızı teorisi ile uyumlu bir hipoteze göre bu kış uykusunda iken metabolizma hızlarını yavaşlatmaları olgusuna bağlanır; kış uykusuna yatan yarasalar ortalama olarak yatmayan yarasalardan daha uzun ömre sahiptir.[163][164] Bir başka hipoteze göre de uçma yeteneklerinin ölüm oranlarını azaltmasıdır ki bu kuşlar ve süzülen memeliler için de doğru olabilir. Birden fazla yavrulayan yarasa türlerinin ömrü genellikle tek yavru yavrulayan yarasa türlerinden daha kısadır. Mağaralara tüneyen yarasaların ömürleri de mağaralarda avlanma riskinin daha az olması nedeniyle tünemeyenlere göre daha uzun olabilir. Doğal yaşam ortamında 41 yıl sonra tekrar yakalanan bir erkek Myotis brandti türü yarasa kaydedilmiş en uzun süre yaşayan yarasadır.[164][165]
Filojeni ve taksonomi

Evrimi
Yarasaların hassas iskeletleri çok iyi fosilleşmez ve fosil kayıtlarında yaşamış olan yarasa cinslerinin yalnızca %12'sinin bulunduğu tahmin edilmektedir.[166] 32 milyon yıl önceden kalma Archaeopteropus gibi bilinen en eski yarasa fosilleri günümüz küçük yarasalarına çok benzerdir.[167] Soyu tükenmiş Palaeochiropteryx tupaiodon (48 milyon yıl önce) ve Hassianycteris kumari (55 milyon yıl önce) renklerinin tespit edilebildiği ilk memeli fosilleridir ve ikisinin de kızılımsı kahverengi olduğu bulunmuştur.[168][169]
Yarasalar önceleri sivri sincapçıklar (Scandentia), uçar makigiller (Dermoptera) ve primatlarla Archonta üst takımında sınıflandırılmışlardı.[170] Modern genetik kanıtlar artık yarasaları içinde etçilleri, pangolinleri, tek toynaklıları, çift toynaklıları ve balinaları barındıran kardeş taksonu Ferungulata ile birlikte Laurasiatheria üst takımında sınıflandırılmasına neden olmuştur.[171][172][173][174][175] Bir çalışma da Chiroptera takımını tek toynaklılar (Perissodactyla) ile kardeş takson olarak sınıflandırır.[176]
| |||||||||||||||||||||||||||||||||||||||
| 2013 yılında yapılmış bir çalışmaya göre Chiroptera kardeş taksonu Ferungulata ile Laurasiatheria içinde gösterilmektedir.[175] |



Farklı yarasa gruplarının filogenetik ilişkileri çok büyük bir tartışma konusudur. Geleneksel Megachiroptera ve Microchiroptera ayrımı yarasaların zaten uçma yeteneğine sahip bir ortak atadan geldiğini ama çok uzun zamandır birbirlerinden bağımsız olarak evrimleştikleri görüşünü yansıtmaktaydı. Bu hipotez küçük ve büyük yarasalar arasındaki farklılıkları tanımakta ve uçma yeteneğinin memelilerde yalnızca bir kere geliştiğini kabul etmekteydi. Moleküler biyolojik kanıtların çoğu yarasaların doğal ya da monofiletik bir grup oluşturduğu yönündedir.[167]
|


Genetik kanıtlar büyük yarasaların Eosen'in başlarında küçük yarasaların dört ana soyuyla birlikte ortaya çıktığını göstermektedir.[175] Pteropodidae ya da büyük yarasalar familyası ile birlikte küçük yarasaların Rhinolophidae, Hipposideridae, Craseonycteridae, Megadermatidae ve Rhinopomatidae familyalarını da içeren Yinpterochiroptera[177] ile hepsi gırtlak ekolokasyonu kullanan diğer yarasa familyalarını içeren Yangochiroptera olmak üzere iki yeni alt takım 2005 yılında yapılan bir DNA araştırmasının sonuçlarıyla desteklenerek önerilmiştir.[177] 2013 yılında yapılan bir filojenomik araştırma da önerilen iki alt takımı destekleyen yönde bulgulara ulaşmıştır.[175]
|

1980'lerde morfolojik bulgulara dayanan bir hipotez Megachiroptera alt takımının uçma yeteneğinin Microchiroptera alt takımından ayrı olarak evrimleştirdiğini öne sürmüştür. Uçan primat hipotezi uçma adaptasyonu çıkarıldığında Megachiroptera'nın Microchiroptera ile paylaşmadıkları anatomik özellikleri ile primatlarla bağlantılı olduklarını önermektedir. Örneğin büyük yarasaların beyinleri gelişmiş özelliklere sahiptir. Her ne kadar son zamanlarda yapılan genetik araştırmalar yarasaların monofilik olduğunu desteklese de[167] genetik ve morfolojik bulguların anlamı üzerine olan tartışmalar sürmektedir.[178]
52 milyon yaşında olan Green River Oluşumu'nda 2003 yılında Onychonycteris finneyi fosilinin bulunması uçma yeteneğinin ekolokasyon yeteneğinin evrimleşmesinden daha önce kazanıldığını göstermektedir.[179][180] Günümüz yarasalarının her iki elinde en fazla iki parmağında iki pençe olmasına rağmen Onychonycteris fosilinin beş parmağının hepsinde pençe bulunmaktadır. Ayrıca tembel hayvanlar ve gibonlar gibi dallarından altından sallanan tırmanıcı memeliler benzer şekilde arka ayakları daha uzun ve önayakları da daha kısadır. Bu avuç içi büyüklüğündeki yarasanın kısa ve geniş kanatları daha sonra gelen yarasa türleri kadar hızlı ve uzağa uçamayabileceğini göstermektedir. Onychonycteris uçarken kanatlarını sürekli çırpmak yerine muhtemelen havada kanat çırpma ve süzülme hareketlerini değişimli olarak kullanıyordu.[167] Bu bulgular da bu yarasanın günümüz yarasaları kadar çok uçmadığını ancak ağaçtan ağaca uçtuğunu ve zamanının çoğunu tırmanarak ya da dallardan sarkarak geçirdiğini önermektedir.[181] Onychonycteris fosilinin ayırt edici özellikleri aynı zamanda memeli uçuşunun yerde hareket eden memelilerden çok ağaçlara tırmanan memelilerden geliştiği hipotezini de destekler. Genel olarak "ağaçtan aşağı" olarak bilinen bu uçma gelişimi modelinde uçabilmek için yerde hızlı koşmak yerine yarasaların avlarının üzerine çullanmak için yükseklik ve yerçekimi avantajını kullanarak uçtuklarını önerir.[182][183]
Küçük yarasaların tek bir ortak atası olmadığına işaret eden ve olması düşük ihtimalli dönüşümlerin olduğunu gösteren moleküler filojeni tartışmalıdır. Bunlardan ilki gırtlak ekolokasyonunun yarasalarda iki kere evrimleşmiş olması, bir kere Yangochiroptera ve bir kere de rhinolophoidlerde.[184] İkincisi de gırtlak ekolokasyonunun Chiroptera takımında tek bir kaynağı olduğu ve bunun daha sonra büyük yarasaları oluşturan Pteropodidae familyasında kaybolduğu ve sonradan Rousettus cinsinde ise bir çeşit dil şaklatması sistemine evrimleşmesidir.[185] Seslendirme geni FoxP2 dizisinin analizi gırtlak ekolokasyonunun pteropoidlerde kaybolduğu ya da ekolokasyon kullanan soylarda kazanıldığı yönünde görüş bilrdirmek için sonuç vermemiştir.[186] Yarasalarda ekolokasyon muhtemelen ilk olarak iletişim çağrılarından gelişmiştir. Eosen yarasaları Icaronycteris (52 milyon yıl önce) ve Palaeochiropteryx ultrasonu algılayacak kafatası adaptasyonları göstermektedir. Bunlar ilk önceleri yerde böcekleri avlamak, süzülürken etraflarının haritasını çıkarabilmek ya da iletişim amaçları için kullanılmış olabilir. Uçma adaptasyonu yerleştikten sonra da uçan avların yerlerini ekolokasyon ile belirleyebilmek için daha gelişmiş olabilir.[181] Yarasaların ekolokasyonu tek bir ortak atadan evrimleşmiş olabilir ki bu durumda Eski Dünya büyük yarasaları bunu daha sonra kaybetmiş ve yalnızca nalburunlu yarasalar tekrar kazanmıştır ya da ekolokasyon Yinpterochiroptera ve Yangochiroptera soylarında bağımsız olarak evrimleşmiştir.[187] İşitme geni Prestin'in analizi pteroidlerde kaybolmasından ekolokasyonun bağımsız olarak en az iki kere geliştiği fikrini destekler görünmektedir[188] ancak kokleanın ontojenik analizi de gırtlak ekolokasyonunun yalnızca bir kere evrimleştiğini desteklemektedir.[189]
Sınıflandırma

Yarasalar plasentalı memelilerdir. Kemiricilerden sonra memeli türlerinin yaklaşık %20'sini oluşturan en büyük memeli takımıdır.[190] Carl Linnaeus 1758 yılında bildiği yedi yarasa türünü Primatlar takımında Vespertilio cinsi içinde sınıflandırdı. Yaklaşık yirmi yıl sonra Alman doğabilimci Johann Friedrich Blumenbach yarasaları kendi takımları Chiroptera içine aldı.[191] O zamandan beri tanımlanan tür sayısı 1.200'ün üzerine çıktı ve geleneksel olarak iki alt takımda sınıflandırıldılar: Megachiroptera (büyük yarasalar) ve Microchiroptera (küçük yarasalar/ekolokasyon kullanan yarasalar).[192] Büyük yarasaların hepsi küçük yarasalardan daha büyük değildir.[193] İki grubu çeşitli özellikler ayırır. Küçük yarasalar yollarını bulmak ve avlanmak için ekolokasyon kullanırken Rousettus cinsi haricindeki büyük yarasalar bunlar için yalnızca gözlerini kullanırlar.[194] Buna bağlı olarak da büyük yarasaların görsel korteksleri iyi gelişmiştir ve iyi bir görme keskinlikleri vardır.[192] Büyük yarasaların önayaklarının ikinci parmağında bir pençeleri olur.[195][196] Küçük yarasaların dış kulakları bir halka oluşturacak şekilde kapanmaz, kenarlar kulağın tabanında birbirlerinden ayrılır.[196] Büyük yarasalar meyve, nektar ya da polen yerken küçük yarasaların çoğu böcekçildir ve diğerleri de meyve, nektar, polen, balık, kurbağa, küçük memeliler ya da kan ile beslenirler.[192]
Aşağıda 2019 yılında basılan Handbook of the Mammals of the World'ün (Dünya Memeliler Elkitabı) dokuzuncu baskısında çeşitli yazarlar tarafından tanınan Chiroptera takımının familyalarının sınıflandırılması verilmektedir:[197]
| Chiroptera Blumenbach, 1779 | |||
| Yinpterochiroptera Springer, Teeling, Madsen, Stanhope & Jong, 2001 | |||
| Pteropodoidea J. E. Gray, 1821 | |||
| Familya | Açıklama | Tür sayısı | Görsel |
|---|---|---|---|
| Pteropodidae J. E. Gray, 1821 | Eski Dünya meyve yarasaları | 191 |  |
| Rhinolophoidea J. E. Gray, 1825 | |||
| Familya | Açıklama | Tür sayısı | Görsel |
| Rhinopomatidae Bonaparte, 1838 | Fare kuyruklu yarasalar | 6 |  |
| Craseonycteridae Hill, 1974 | Yabanarısı yarasası | 1 |  |
| Megadermatidae H. Allen, 1864 | Yalancı vampir yarasalar | 6 |  |
| Rhinonycteridae J. E. Gray, 1866 | Üç çatallı zıpkın yarasaları | 9 |  |
| Hipposideridae Lydekker, 1891 | Eski Dünya yaprak burunlu yarasalar | 88 |  |
| Rhinolophidae J. E. Gray, 1825 | Nalburunlu yarasalar | 109 |  |
| Yangochiroptera Koopman, 1984 | |||
| Emballonuroidea Gervais in de Castelnau, 1855 | |||
| Familya | Açıklama | Tür sayısı | Görsel |
| Nycteridae Van der Hoeven, 1855 | Yarık yüzlü yarasalar | 15 |  |
| Emballonuridae Gervais in de Castelnau, 1855 | Kın kuyruklu yarasalar | 54 |  |
| Noctilionoidea J. E. Gray, 1821 | |||
| Familya | Açıklama | Tür sayısı | Görsel |
| Myzopodidae Thomas, 1904 | Madagaskar vantuz ayaklı yarasalar | 2 |  |
| Mystacinidae Dobson, 1875 | Yeni Zelanda kısa kuyruklu yarasalar | 2 |  |
| Thyropteridae Miller, 1907 | Disk kanatlı yarasalar | 5 |  |
| Furipteridae J. E. Gray, 1866 | 2 |  | |
| Noctilionidae J. E. Gray, 1821 | Buldog yarasalar | 2 |  |
| Mormoopidae Saussure, 1860 | Hayalet suratlı, çıplak sırtlı ve bıyıklı yarasalar | 18 |  |
| Phyllostomidae J. E. Gray, 1825 | Yeni Dünya yaprak burunlu yarasalar | 217 |  |
| Vespertilionoidea J. E. Gray, 1821 | |||
| Familya | Açıklama | Tür sayısı | Görsel |
| Natalidae J. E. Gray, 1825 | Huni kulaklı yarasalar | 12 |  |
| Molossidae Gervais in de Castelnau, 1855 | Serbest kuyruklu yarasalar | 126 |  |
| Miniopteridae Dobson, 1875 | Serbest kuyruklu yarasalar | 38 |  |
| Cistugidae Lack et al., 2010 | Kanat bezli yarasalar | 2 | |
| Vespertilionidae J. E. Gray, 1821 | Vesper yarasaları | 496 |  |
İnsanlarla ilişkileri
Korunma

Bat Conservation International[198] gibi gruplar yarasaların ekolojik rolleri ve karşılaştıkları çevresel tehditler üzerine olan farkındalığı artırmayı amaçlamaktadır. Birleşik Krallık'ta tüm yarasalar 1981 yılında çıkarılmış "Yaban Hayatı ve Kırsal Alanlar Kanunu" ile korunmaktadır ve bir yarasayı ya da tüneğini rahatsız etmek ağır bir para cezası ile cezalandırılabilir.[199] Malezya Saravak'ta "tüm yarasalar[200] 1998 yılında çıkarılmış "Malezya Yaban Hayatı Kanunu" ile koruma altındadır[200] ama Cheiromeles torquatus gibi yarasa türleri hâlâ yerel halk tarafından yenilmektedir.[201] Modern dönemde insanlar çeşitli yarasa türlerinin soyunun tükenmesine neden olmuştur, bunların en sonuncusu 2009 yılında soyunun tükendiği bildirilen Pipistrellus murrayi türü yarasadır.[202]
Birçok kişi yarasaları çekmek için yarasa barınakları yapmaktadır.[203] Yaklaşık 400.000 yarasa ile Florida Üniversitesi'nin 1991 yılında kurulan yarasa barınağı dünya üzerinde en kalabalık yapay tünek yeridir.[204] Britanya'da II. Dünya Savaşı'ndan kalma kalın duvarlı ve kısmen yeraltında olan savunma mevzileri yarasalar için tünek hâline getirilmiştir.[205][206] Yol yapımı ve diğer kentsel gelişim nedenleriyle habitata verilen zararları hafifletmek amacıyla özel olarak yarasa barınakları da inşa edilmektedir.[207][208] Hassas ve tehdit altında olan yarasa türlerinin yaşadığı mağaralara insanların girişini kısıtlamak için bazen mağara girişlerine özel kapılar konulmaktadır. Bu kapılar hava akışını bozmayacak ve yarasaların girişi çıkışına engel olmayacak şekilde tasarlanmıştır ve dolayısıyla mağaranın içindeki mikro ekosistem koruma altına alınır.[209] ABD'de bulunan 47 yarasa türünden 35'inin binalar ve köprüler de dahil olmak üzere insan yapısı binaları kullandıkları bilinmektedir. On dört tür yapay yarasa barınaklarını kullanır.[210]
Yarasalar Afrika, Asya ve Pasifik Kıyılarında besin olarak tüketilir. Guam'da olduğu gibi bazı durumlarda besin olarak avlandıkları için soyları tehdit altına girmiştir.[211] Rüzgâr türbinlerinin yarasaları öldürecek kadar barotravma yarattığına dair kanıt bulunmaktadır.[212] Yarasaların tipik memeli akciğerlerinin ani hava basıncı değişikliklerine kuş akciğerlerinden daha hassas olduğu ve dolayısıyla ölümcül akciğer yarılmalarına neden olduğu düşünülmektedir.[213][214][215][216][217] Yarasalar muhtemelen tünek yeri aramak için rüzgâr türbinlerine yaklaşmakta bu da ölüm oranını artırmaktadır.[213] Akustik uzaklaştırıcılar rüzgâr türbinlerindeki yarasa ölüm oranını azaltmaya yardımcı olabilmektedir.[218]
Kültürel önemi
- Erbâb-ı kemâli çekemez nakıs olanlar
- Rencide olur dîde-i huffâş ziyadan
- ...


Yarasalar memeli olmasına rağmen uçabildikleri için çeşitli geleneklerde liminal varlıklar olarak görülürler.[219] Birçok kültürde yarasalara karanlık, ölüm, cadılık ile büyücülük ve kötü niyetlilik ile bağdaştırılır.[220] Muskogi, Çeroki ve Apaçi halkları gibi Amerikan yerlileri arasında yarasa hilekâr ruhtur. Tanzanya'da Popobawa olarak bilinen kanatlı yarasa şeklinde bir varlığın kurbanlarına saldırarak sodomize eden, şekil değiştiren bir kötü ruh olduğuna inanılır.[221] Aztek mitolojisinde yarasalar ölüler diyarını, yıkımı ve çürümeyi temsil ederler.[222][223][224] Bir Doğu Nijerya masalında yarasanın eşi çalı faresinin ölümüne neden olduktan sonra gece yaşama alışkanlığına başladığını ve gündüzleri yakalanmamak için saklandığını anlatır.[225]
Bazı kültürlerde yarasaların daha olumlu tasvirleri de bulunur. Çin'de yarasalar mutluluk, neş'e ve iyi talihle bağdaştırılır. Beş yarasa "Beş Lütuf"un sembolü olarak kullanılır: uzun yaşam, refah, sağlık, erdem aşkı ve huzurlu ölüm.[226] Yarasa Tonga'da kutsaldır ve genellikle vücuttan ayrılabilen ruhun fiziksel görünümü olarak kabu ledilir.[227] Orta Amerika'da Zapotek uygarlığında yarasa tanrı mısır ve bereket tanrısıydı.[228]
William Shakespeare'in Macbeth oyununda Üç Cadı iksirlerinde yarasa kürkü kullanırlar.[229] Batı Kültüründe yarasa gecenin ve geceleyin olan kötülüklerin bir sembolüdür. Yarasa gecenin kurgusal karakterlei ile bağdaştırılan ana hayvanlardan biridir: Kont Drakula gibi kötü adamlar[230] ya da Batman gibi kahramanlar.[231]
Yarasa İspanya ve Fransa'da armacılıkta kullanılmıştır ve Valensiya, Palma de Mallorca, Fraga, Albacete ile Montchauvet şehirlerinin armalarında yer almaktadır.[232][233][234] ABD'nin üç eyaletinin resmî eyalet yarasası bulunur: Teksas ve Oklahoma için Tadarida brasiliensis ve Virginia için Corynorhinus townsendii virginianus.[235]
Ekonomik önemi
Tarımsal haşere popülasyonlarını kontrol altında tutmaya ve böylece pestisitlerin kullanılma gerekliliğini azaltmaya yararı olan böcekçil yarasalar çiftçilere yardımcı olmaktadır. ABD'de pestisit kullanımı ve ürünlere gelen zararın önlenmesiyle yarasaların yılda 3,7 ila 53 milyar dolar arasında tasarrufa neden oldukları tahmin edilmektedir. Bu aynı zamanda çevreyi kirletecek pestisitlerin aşırı kullanımı ile gelecek nesil böceklerin bunlara direnç kazanmalarının önlenmesini de içerir.[236]
Bir çeşit guano olan yarasa dışkısı nitrat açısından zengindir ve mağaralardan toplanarak gübre olarak kullanılır.[237] Amerikan İç Savaşı sırasında mağaralardan toplanan güherçile karabarut yapımında kullanılmıştır; bunun yarasa guanosu olduğuna inanılmış olsa da nitratın çoğu azotlayıcı bakterilerden gelmektedir.[238]
Yaz aylarında yaklaşık 1.500.000 kadar Tadarida brasiliensis türü yarasaya ev sahipliği yapan Teksas'ın Austin şehrinde bulunan Congress Avenue Bridge (Kongre Bulvarı Köprüsü) Kuzey Amerika'nın bilinen en büyük kentsel yarasa tüneğidir. Her yıl 100.000 kadar turist alacakaranlıkta yarasaların tüneklerinden ayrılmalarını izlemek için köprüyü ziyaret eder.[239]
Notlar
- ^ Latince Chiroptera'nın Türkçe okunuşu "Kiroptera"dır ve Antik Yunanca "el" anlamına gelen χείρ ile "kanat" anlamına gelen πτερόν kelimelerinin birleştirilmesinden oluşturulmuştur.[3]
- ^ Daha önce yayınlanan ve yalnızca meyvecil yarasalarda bu eksikliğin görüldüğünü belirten raporlar daha küçük örneklemler kullanmıştır.[94]
Kaynakça
Özel
- ^ "Chiroptera". paleobiodb.org. 22 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 22 Ağustos 2021.
- ^ Hutcheon, J.M.; Kirsch, J.A.W. (2006). "A moveable face: Deconstructing the Microchiroptera and a new classification of extant bats". Acta Chiropterologica. 8 (1): 1–10. doi:10.3161/1733-5329(2006)8[1:AMFDTM]2.0.CO;2. ISSN 1733-5329.
- ^ "Chiroptera". Encyclopædia Britannica. 6. ss. 239-247.
- ^ Atalay, Besim (1998). Divanü Lugat-it-Türk Tercümesi. 3. Türk Dil Kurumu. s. 433.
- ^ Gülensoy, Tuncer (2007). Türkiye Türkçesindeki Türkçe Sözcüklerin Köken Bilgisi I -II. Türk Dil Kurumu. ss. 1071-1072.
- ^ "yarasa". Nişanyan Sözlük. 23 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 15 Şubat 2020.
- ^ Fleming, T. (2003). A Bat Man in the Tropics: Chasing El Duende. University of California Press. s. 165. ISBN 978-0520236066.
- ^ a b c d e f g h i j k l m n Jones, G. (2001). "Bats". MacDonald, D. (Ed.). The Encyclopedia of Mammals (2. bas.). Oxford University Press. ss. 754-775. ISBN 978-0-7607-1969-5.
- ^ a b Greenhall, A.M.; Joermann, G.; Schmidt, U. (1983). "Desmodus rotundus". Mammalian Species. 202 (202). ss. 1-6. doi:10.2307/3503895. JSTOR 3503895.
- ^ Senawi, J.; Schmieder, D.; Siemers, B.; Kingston, T. (2015). "Beyond size – morphological predictors of bite force in a diverse insectivorous bat assemblage from Malaysia". Functional Ecology. 29 (11). ss. 1411-1420. doi:10.1111/1365-2435.12447.
- ^ a b Hunter, P. (2007). "The nature of flight: The molecules and mechanics of flight in animals". Science and Society. 8 (9). ss. 811-813. doi:10.1038/sj.embor.7401050. PMC 1973956 $2. PMID 17767190.
- ^ McCracken, G. F.; Safi, K.; Kunz, T. H.; Dechmann, D. K. N.; Swartz, S. M.; Wikelski, M. (9 Kasım 2016). "Airplane tracking documents the fastest flight speeds recorded for bats". Royal Society Open Science. 3 (11). s. 160398. Bibcode:2016RSOS....360398M. doi:10.1098/rsos.160398. PMC 5180116 $2. PMID 28018618.
- ^ Sears, K. E.; Behringer, R. R.; Rasweiler, J. J.; Niswander, L. A. (2006). "Development of bat flight: Morphologic and molecular evolution of bat wing digits". Proceedings of the National Academy of Sciences. 103 (17). ss. 6581-6586. Bibcode:2006PNAS..103.6581S. doi:10.1073/pnas.0509716103. PMC 1458926 $2. PMID 16618938.
- ^ Kirkpatrick, S. J. (1994). "Scale effects on the stresses and safety factors in the wing bones of birds and bats". Journal of Experimental Biology. Cilt 190. ss. 195-215. PMID 7964391.
- ^ a b c d e Pennycuick, C. J. (2008). "Bats". Modelling the Flying Bird. Elsevier. ss. 136-143. ISBN 978-0-12-374299-5.
- ^ a b Marshall, K. L.; Chadha, M.; deSouza, L. A.; Sterbing-D'Angelo, S. J.; Moss, C. F.; Lumpkin, E. A. (2015). "Somatosensory substrates of flight control in bats". Cell Reports. 11 (6). ss. 851-858. doi:10.1016/j.celrep.2015.04.001. PMC 4643944 $2. PMID 25937277.
- ^ Brown University (2007). "Bats In Flight Reveal Unexpected Aerodynamics". ScienceDaily. 19 Aralık 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 31 Ekim 2017.
- ^ Riskin, D. K.; Bergou, A.; Breuer, K. S.; Swartz, S. M. (2012). "Upstroke wing flexion and the inertial cost of bat flight". Proceedings of the Royal Society B: Biological Sciences. 279 (1740). ss. 2945-2950. doi:10.1098/rspb.2012.0346. PMC 3385481 $2. PMID 22496186.
- ^ a b Roberts, W. C. (2006). "Facts and ideas from anywhere". Proceedings (Baylor University. Medical Center). 19 (4). ss. 425-434. doi:10.1080/08998280.2006.11928217. PMC 1618737 $2. PMID 17106509.
- ^ Irwin, N. (1997). "Wanted DNA samples from Nyctimene or Paranyctimene Bats" (PDF). The New Guinea Tropical Ecology and Biodiversity Digest. Cilt 3. s. 10. 22 Temmuz 2008 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 14 Şubat 2020.
- ^ a b Sterbing-D'Angelo, S.; Chadha, M.; Chiu, C.; Falk, B.; Xian, W.; Barcelo, J.; Zook, J. M.; Moss, C. F. (2011). "Bat wing sensors support flight control". Proceedings of the National Academy of Sciences of the United States of America. 108 (27). ss. 11291-11296. Bibcode:2011PNAS..10811291S. doi:10.1073/pnas.1018740108. PMC 3131348 $2. PMID 21690408.
- ^ Mehlhorn, H. (2013). Bats (Chiroptera) as Vectors of Diseases and Parasites: Facts and Myths. Springer. ss. 2-27. ISBN 978-3-642-39333-4.
- ^ a b c Makanya, A. N.; Mortola, J. P. (2017). "The structural design of the bat wing web and its possible role in gas exchange". Journal of Anatomy. 211 (6). ss. 687-697. doi:10.1111/j.1469-7580.2007.00817.x. PMC 2375846 $2. PMID 17971117.
- ^ a b c d e f g h i j k l m n o p q r s t u Fenton & Simmons 2015.
- ^ "Leading Edge Vortex Allows Bats to Stay Aloft, Aerospace Professor Reports". USC Viterbi School of Engineering. 29 Şubat 2008. 20 Ekim 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Şubat 2020.
- ^ a b Fenton, M. B.; Crerar, L. M. (1984). "Cervical Vertebrae in Relation to Roosting Posture in Bats". Journal of Mammalogy. 65 (3). ss. 395-403. doi:10.2307/1381085. JSTOR 1381085.
- ^ Riskin, D. K.; Parsons, S.; Schutt, W. A., Jr.; Carter, G. G.; Hermanson, J. W. (2006). "Terrestrial locomotion of the New Zealand short-tailed bat Mystacina tuberculata and the common vampire bat Desmodus rotundus" (PDF). Journal of Experimental Biology. 209 (9). ss. 1725-1736. doi:10.1242/jeb.02186. PMID 16621953. 28 Eylül 2017 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 14 Şubat 2020.
- ^ Jones, T. W. (1852). "Discovery That the Veins of the Bat's Wing (Which are Furnished with Valves) are Endowed with Rythmical Contractility, and That the Onward Flow of Blood is Accelerated by Each Contraction". Philosophical Transactions of the Royal Society of London. Cilt 142. ss. 131-136. doi:10.1098/rstl.1852.0011. JSTOR 108539.
- ^ Dongaonkar, R. M.; Quick, C. M.; Vo, J. C.; Meisner, J. K.; Laine, G. A.; Davis, M. J.; Stewart, R. H. (15 Haziran 2012). "Blood flow augmentation by intrinsic venular contraction in vivo". American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 302 (12). ss. R1436-R1442. doi:10.1152/ajpregu.00635.2011. PMC 3378342 $2. PMID 22513742.
- ^ Langley, L. (29 Ağustos 2015). "Bats and Sloths Don't Get Dizzy Hanging Upside Down—Here's Why". National Geographic. 2 Nisan 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Haziran 2017.
- ^ Maina, J. N. (2000). "What it takes to fly: the structural and functional respiratory refinements in birds and bats". Journal of Experimental Biology. 203 (20). ss. 3045-3064. PMID 11003817. 18 Eylül 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Şubat 2020.
- ^ Suthers, Roderick A.; Thomas, Steven P; Suthers, Barbara A (1972). "Respiration, Wing-Beat and Ultrasonic Pulse Emission in an Echo-Locating Bat". Journal of Experimental Biology. 56 (56). ss. 37-48. 9 Ağustos 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Ağustos 2019.
- ^ a b Ben-Hamo, Miriam; Muñoz-Garcia, Agustí; Larrain, Paloma; Pinshow, Berry; Korine, Carmi; Williams, Joseph B. (29 Haziran 2016). "The cutaneous lipid composition of bat wing and tail membranes: a case of convergent evolution with birds". Proceedings of the Royal Society B: Biological Sciences. 283 (1833). s. 20160636. doi:10.1098/rspb.2016.0636. PMC 4936036 $2. PMID 27335420.
- ^ Jürgens, Klaus Dieter; Bartels, Heinz; Bartels, Rut (1981). "Blood oxygen transport and organ weights of small bats and small non-flying mammals". Respiration Physiology. 45 (3). ss. 243-260. doi:10.1016/0034-5687(81)90009-8. PMID 7330485.
- ^ Martini, Frederic (2015). Visual anatomy & physiology. Pearson. ss. 704-705. ISBN 978-0-321-91874-1. OCLC 857980151.
- ^ Wang, Li; Li, Gang; Wang, Jinhong; Ye, Shaohui; Jones, Gareth; Zhang, Shuyi (2009). "Molecular cloning and evolutionary analysis of the GJA1 (connexin43) gene from bats (Chiroptera)". Genetics Research. 91 (2). ss. 101-9. doi:10.1017/s0016672309000032. PMID 19393126.
- ^ Holbrook, K. A.; Odland, G. F. (1978). "A collagen and elastic network in the wing of the bat". Journal of Anatomy. 126 (Pt 1). ss. 21-36. PMC 1235709 $2. PMID 649500.
- ^ Strobel, S.; Roswag, A.; Becker, N. I.; Trenczek, T. E.; Encarnação, J. A. (2013). "Insectivorous Bats Digest Chitin in the Stomach Using Acidic Mammalian Chitinase". PLOS ONE. 8 (9). ss. e72770. Bibcode:2013PLoSO...872770S. doi:10.1371/journal.pone.0072770. PMC 3760910 $2. PMID 24019876.
- ^ a b Schondube, J. E.; Herrera-M, L. Gerardo; Martínez del Rio, C. (2001). "Diet and the evolution of digestion and renal function in phyllostomid bats" (PDF). Zoology. 104 (1). ss. 59-73. doi:10.1078/0944-2006-00007. PMID 16351819. 11 Ağustos 2017 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 14 Şubat 2020.
- ^ Lyons, Rachel; Wimberley, Trish (Mart 2014). Introduction to the Care and Rehabilitation of Microbats (PDF). 3.0. Wildcare Australia. s. 12. 10 Mart 2018 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 14 Şubat 2020.
- ^ Fenton & Simmons 2015, s. 164.
- ^ Suthers, Roderick; Fattu, James (1973). "Mechanisms of Sound Production by Echolocating Bats". American Zoologist. 13 (4). ss. 1215-1226. doi:10.1093/icb/13.4.1215.
- ^ Jones, K. E.; Bininda-Emonds, O. R. P.; Gittleman, J. L. (2005). "Bats, clocks, and rocks: diversification patterns in chiroptera". Evolution. 59 (10). ss. 2243-2255. doi:10.1554/04-635.1. PMID 16405167.
- ^ Surlykke, A.; Elisabeth, K. V. (2008). "Echolocating bats Cry Out Loud to Detect Their Prey". PLoS ONE. 3 (4). s. e2036. Bibcode:2008PLoSO...3.2036S. doi:10.1371/journal.pone.0002036. PMC 2323577 $2. PMID 18446226.
- ^ a b c Muller, R. (2004). "A numerical study of the role of the tragus in the big brown bat". The Journal of the Acoustical Society of America. 116 (6). ss. 3701-3712. Bibcode:2004ASAJ..116.3701M. doi:10.1121/1.1815133. PMID 15658720.
- ^ Teeling, E. C. (2009). "Hear, hear: the convergent evolution of echolocation in bats?". Trends in Ecology & Evolution. 24 (7). ss. 351-354. doi:10.1016/J.Tree.2009.02.012. PMID 19482373.
- ^ Jones, G.; Holderied, M. W. (2007). "Bat echolocation calls: adaptation and convergent evolution". Proceedings of the Royal Society B: Biological Sciences. 274 (1612). ss. 905-912. doi:10.1098/Rspb.2006.0200. PMC 1919403 $2. PMID 17251105.
- ^ Pavey, C. R.; Burwell, C. J. (1998). "Bat Predation on Eared Moths: A Test of the Allotonic Frequency Hypothesis". Oikos. 81 (1). ss. 143-151. doi:10.2307/3546476. JSTOR 3546476.
- ^ Sowell, W. A. (1983). "The Bat's Ear as a Diffraction Grating" (PDF). Air Force Institute of Technology.[]
- ^ K., Roman (2009). "Model predicts bat pinna ridges focus high frequencies to form narrow sensitivity beams". The Journal of the Acoustical Society of America. 125 (5). ss. 3454-9. Bibcode:2009ASAJ..125.3454K. doi:10.1121/1.3097500. PMID 19425684.
- ^ a b Corcoran, A. J.; Barber, J. R.; Conner, W. E. (2009). "Tiger moth jams bat sonar". Science. 325 (5938). ss. 325-327. Bibcode:2009Sci...325..325C. doi:10.1126/science.1174096. PMID 19608920.
- ^ a b Hristov, N. I.; Conner, W. E. (2005). "Sound strategy: acoustic aposematism in the bat–tiger moth arms race". Naturwissenschaften. 92 (4). ss. 164-169. Bibcode:2005NW.....92..164H. doi:10.1007/s00114-005-0611-7. PMID 15772807.
- ^ Surlykke, A.; Ghose, K.; Moss, C. F. (2009). "Acoustic scanning of natural scenes by echolocation in the big brown bat, Eptesicus fuscus". Journal of Experimental Biology. 212 (Pt 7). ss. 1011-20. doi:10.1242/jeb.024620. PMC 2726860 $2. PMID 19282498.
- ^ Strauß, J.; Lakes-Harlan, R. (2014). "Evolutionary and Phylogenetic Origins of Tympanal Hearing Organs in Insects". Hedwig, B. (Ed.). Insect Hearing and Acoustic Communication. Springer. ss. 5-26. doi:10.1007/978-3-642-40462-7_2. ISBN 978-3-642-40462-7.
- ^ Fullard, J. H. (1998). "Moth Ears and Bat Calls: Coevolution or Coincidence?". Hoy, R. R.; Fay, R. R.; Popper, A. N. (Ed.). Comparative Hearing: Insects. Springer Handbook of Auditory Research. Springer. ISBN 978-1-4612-6828-4.
- ^ Takanashi, Takuma; Nakano, Ryo; Surlykke, A.; Tatsuta, H.; Tabata, J.; Ishikawa, Y.; Skals, N. (2010). "Variation in Courtship Ultrasounds of Three Ostrinia Moths with Different Sex Pheromones". PLOS ONE. 5 (10). ss. e13144. Bibcode:2010PLoSO...513144T. doi:10.1371/journal.pone.0013144. PMC 2949388 $2. PMID 20957230.
- ^ Sophasarun, N. "Experts debunk bats' bad rap". Online extra. National Geographic. 26 Ocak 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Kasım 2017.
- ^ Müller, B.; Glösmann, M.; Peichl, L.; Knop, G. C.; Hagemann, C.; Ammermüller, J. (2009). "Bat Eyes Have Ultraviolet-Sensitive Cone Photoreceptors". PLOS ONE. 4 (7). ss. e6390. Bibcode:2009PLoSO...4.6390M. doi:10.1371/journal.pone.0006390. PMC 2712075 $2. PMID 19636375.
- ^ Shen, Y.-Y.; Liu, J.; Irwin, D. M.; Zhang, Y.-P. (2010). "Parallel and Convergent Evolution of the Dim-Light Vision Gene RH1 in Bats (Order: Chiroptera)". PLOS ONE. 5 (1). ss. e8838. Bibcode:2010PLoSO...5.8838S. doi:10.1371/journal.pone.0008838. PMC 2809114 $2. PMID 20098620.
- ^ a b Wang, D.; Oakley, T.; Mower, J.; Shimmin, L. C.; Yim, S.; Honeycutt, R. L.; Tsao, H.; Li, W. H. (2004). "Molecular evolution of bat color vision genes". Molecular Biology and Evolution. 21 (2). ss. 295-302. doi:10.1093/molbev/msh015. PMID 14660703.
- ^ Wang, Y.; Pan, Y.; Parsons, S.; Walker, M.; Zhang, S. (2007). "Bats Respond to Polarity of a Magnetic Field". Proceedings of the Royal Society B: Biological Sciences. 274 (1627). ss. 2901-2905. doi:10.1098/rspb.2007.0904. PMC 2288691 $2. PMID 17848365.
- ^ Tian, L.-X.; Pan, Y.-X.; Metzner, W.; Zhang, J.-S.; Zhang, B.-F. (2015). "Bats Respond to Very Weak Magnetic Fields". PLOS ONE. 10 (4). ss. e0123205. Bibcode:2015PLoSO..1023205T. doi:10.1371/journal.pone.0123205. PMC 4414586 $2. PMID 25922944.
- ^ Nowack, J.; Stawski, C.; Geiser, F. (2017). "More functions of torpor and their roles in a changing world". Journal of Comparative Physiology B. 187 (5–6). ss. 889-897. doi:10.1007/s00360-017-1100-y. PMC 5486538 $2. PMID 28432393.
- ^ Altringham 2011, ss. 99–100.
- ^ Voigt, C. C.; Lewanzik, D. (2011). "Trapped in the darkness of the night: thermal and energetic constraints of daylight flight in bats". Proceedings of the Royal Society B: Biological Sciences. 278 (1716). ss. 2311-2317. doi:10.1098/rspb.2010.2290. PMC 3119008 $2. PMID 21208959.
- ^ Ochoa-Acuña, H.; Kunz, T.H. (1999). "Thermoregulatory behavior in the small island flying fox, Pteropus hypomelanus (Chiroptera: Pteropodidae)". Journal of Thermal Biology. 24 (1). ss. 15-20. CiteSeerX 10.1.1.581.38 $2. doi:10.1016/S0306-4565(98)00033-3.
- ^ Licht, Paul; Leitner, Philip (1967). "Physiological responses to high environmental temperatures in three species of microchiropteran bats". Comparative Biochemistry and Physiology. 22 (2). ss. 371-387. doi:10.1016/0010-406X(67)90601-9.
- ^ Neuweiler, Gerhard (2000). "The Circulatory and Respiratory Systems". The Biology of Bats. Oxford University Press. ss. 43-62. ISBN 978-0-1950-9951-5.
- ^ Geiser, F.; Stawski, C. (2011). "Hibernation and Torpor in Tropical and Subtropical Bats in Relation to Energetics, Extinctions, and the Evolution of Endothermy". Integrative and Comparative Biology. 51 (3). ss. 337-338. doi:10.1093/icb/icr042. PMID 21700575.
- ^ Stawski, C.; Geiser, F. (2010). "Fat and Fed: Frequent Use of Summer Torpor in a Subtropical Bat". Naturwissenschaften. 97 (1). ss. 29-35. Bibcode:2010NW.....97...29S. doi:10.1007/s00114-009-0606-x. PMID 19756460.
- ^ Zubaid, A.; McCracken, G. F.; Kunz, T. (2006). Functional and Evolutionary Ecology of Bats. Oxford University Press. ss. 14-16. ISBN 978-0-19-515472-6.
- ^ Knight, K. (2012). "Bats Use Torpor to Minimise Costs". Journal of Experimental Biology. 215 (12). ss. iii. doi:10.1242/jeb.074823.
- ^ Bondarenco, A.; Körtner, G.; Geiser, F. (2016). "How to Keep Cool in a Hot Desert: Torpor in Two Species of Free-Ranging Bats in Summer". Temperature. 6 (3). ss. 476-483. doi:10.1080/23328940.2016.1214334. PMC 5079220 $2. PMID 28349087.
- ^ McGuire, L. P.; Jonassen, K. A.; Guglielmo, C. G. (2014). "Bats on a Budget: Torpor-Assisted Migration Saves Time and Energy". PLOS ONE. 9 (12). ss. e115724. Bibcode:2014PLoSO...9k5724M. doi:10.1371/journal.pone.0115724. PMC 4281203 $2. PMID 25551615.
- ^ Hamilton, I. M.; Barclay, R. M. R. (1994). "Patterns of daily torpor and day-roost selection by male and female big brown bats (Eptesicus fuscus)". Canadian Journal of Zoology. 72 (4). s. 744. doi:10.1139/z94-100.
- ^ "Kitti's Hog-Nosed Bat: Craseonycteridae – Physical Characteristics – Bats, Bumblebee, Species, Inches, Brown, and Tips". Animal Life Resource. 15 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Haziran 2013.
- ^ "Bumblebee bat (Craseonycteris thonglongyai)". EDGE Species. 19 Ağustos 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Nisan 2008.
- ^ "Kitti's Hog-Nosed Bat Is World's Smallest Mammal". SciTechDaily. 3 Aralık 2012. 21 Aralık 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 1 Kasım 2017.
- ^ Nowak, R. M., editor (1999). Walker's Mammals of the World. Vol. 1. 6th edition. Pp. 264–271. 0-8018-5789-9
- ^ a b Gonsalves, L.; Bicknell, B.; Law, B.; Webb, C.; Monamy, V. (2013). "Mosquito Consumption by Insectivorous Bats: Does Size Matter?". PLOS ONE. 8 (10). ss. e77183. Bibcode:2013PLoSO...877183G. doi:10.1371/journal.pone.0077183. PMC 3795000 $2. PMID 24130851.
- ^ Dechmann, D. K. N.; Safi, K.; Vonhof, M. J. (2006). "Matching Morphology and Diet in the Disc-Winged Bat Thyroptera tricolor (Chiroptera)". Journal of Mammalogy. 87 (5). ss. 1013-1019. Bibcode:2007JMamm..88..275L. doi:10.1644/05-MAMM-A-424R2.1.
- ^ Thomas, S. P.; Suthers, R. A. (1972). "Physiology and energetics of bat flight" (PDF). Journal of Experimental Biology. Cilt 57. ss. 317-335. 15 Şubat 2020 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 15 Şubat 2020.
- ^ "Bats of the World". Bat Conservation Trust. 5 Ocak 2011 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Ocak 2011.
- ^ Grzimek's Animal Life Encyclopedia: Vol 13 Mammals II (2. bas.). 2003. s. 311. ISBN 978-0-7876-5362-0.
- ^ Altringham 2011, s. 21.
- ^ "The Art and Science of Bats". Smithsonian Institution. 7 Aralık 2010. 21 Aralık 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 15 Şubat 2020.
- ^ a b Schwab, I. R.; Pettigrew, J. (2005). "A choroidal sleight of hand". British Journal of Ophthalmology. 89 (11). s. 1398. doi:10.1136/bjo.2005.077966. PMC 1772916 $2. PMID 16267906.
- ^ Alexander, D. E. (2015). On the Wing: Insects, Pterosaurs, Birds, Bats and the Evolution of Animal Flight. Oxford University Press. s. 137. ISBN 978-0199996773.
- ^ a b c d e f g Fenton 2001.
- ^ Wolchover, N. (5 Mayıs 2011). "Why Bats Hate Rain". LiveScience. 15 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Aralık 2017.
- ^ Rabe, M. J. (Haziran 1998). "Long Foraging Distance for a Spotted Bat (Euderma Maculatum) in Northern Arizona". The Southwestern Naturalist. 43 (2). ss. 266-269. JSTOR 30055364.
- ^ Cui, J.; Yuan, X.; Wang, L.; Jones, G.; Zhang, S. (2011). "Recent loss of vitamin C biosynthesis ability in bats". PLoS ONE. 6 (11). ss. e27114. Bibcode:2011PLoSO...627114C. doi:10.1371/journal.pone.0027114. PMC 3206078 $2. PMID 22069493.
- ^ Jenness, R.; Birney, E.; Ayaz, K. (1980). "Variation of L-gulonolactone oxidase activity in placental mammals". Comparative Biochemistry and Physiology B. 67 (2). ss. 195-204. doi:10.1016/0305-0491(80)90131-5.
- ^ a b Cui, J.; Pan, Y. H.; Zhang, Y.; Jones, G.; Zhang, S. (2011). "Progressive pseudogenization: vitamin C synthesis and its loss in bats". Mol. Biol. Evol. 28 (2). ss. 1025-31. doi:10.1093/molbev/msq286. PMID 21037206.
- ^ Wray, Amy K.; Jusino, Michelle A.; Banik, Mark T.; Palmer, Jonathan M.; Kaarakka, Heather; White, J. Paul; Lindner, Daniel L.; Gratton, Claudio; Peery, M Zachariah (2018). "Incidence and taxonomic richness of mosquitoes in the diets of little brown and big brown bats". Journal of Mammalogy. 99 (3). ss. 668-674. doi:10.1093/jmammal/gyy044.
- ^ McCracken, G. F.; Gillam, E. H.; Westbrook, J. K.; Lee, Y. F.; Jensen, M. L.; Balsley, B. B. (2008). "Brazilian free-tailed bats (Tadarida brasiliensis: Molossidae, Chiroptera) at high altitude: Links to migratory insect populations". Integrative and Comparative Biology. 48 (1). ss. 107-118. doi:10.1093/icb/icn033. PMID 21669777.
- ^ "Little Brown Bat". Penn State University. 1 Ağustos 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 13 Eylül 2017.
- ^ Fitt, G. P. (1989). "The ecology of Heliothis species in relation to agro-ecosystems". Annual Review of Entomology. Cilt 34. ss. 17-52. doi:10.1146/annurev.ento.34.1.17.
- ^ Boyles, J. G.; McGuire, L. P.; Boyles, E.; Reimer, J. P.; Brooks, C. A.; Rutherford, R. W.; Rutherford, T. A.; Whitaker, J. O., Jr.; McCracken, G. F. (2016). "Physiological and behavioral adaptations in bats living at high latitudes". Physiology and Behavior. Cilt 165. ss. 322-327. doi:10.1016/j.physbeh.2016.08.016. PMID 27542518.
- ^ Simmons, N. B.; Voss, R. S.; Mori, S. A. "Bats as Dispersers of Plants in the Lowland Forests of Central French Guiana". New York Botanical Garden. 11 Ağustos 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Eylül 2017.
- ^ Ortega, J.; Castro-Arellano, I. (2001). "Artibeus jamaicensis". Mammalian Species. Cilt 662. ss. 1-9. doi:10.1644/1545-1410(2001)662<0001:aj>2.0.co;2.
- ^ a b Chamberlain, T. (6 Aralık 2006). "Photo in the News: Bat Has Longest Tongue of Any Mammal". National Geographic News. National Geographic Society. 6 Haziran 2007 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Haziran 2007.
- ^ Arita, H. T.; Santos-Del-Prado, K.; Arita, H.T. (1999). "Conservation Biology of Nectar-Feeding Bats in Mexico". Journal of Mammalogy. 80 (1). ss. 31-41. doi:10.2307/1383205. JSTOR 1383205.
- ^ Gerardo, H.; Hobson, K. A.; Adriana, M. A.; Daniel, E. B.; Sanchez-Corero, V.; German, M. C. (2001). "The Role of Fruits and Insects in the Nutrition of Frugivorous Bats: Evaluating the Use of Stable Isotope Models". Biotropica. 33 (3). ss. 520-528. doi:10.1111/j.1744-7429.2001.tb00206.x.
- ^ Hodgkison, R.; Balding, S. T.; Zuibad, A.; Kunz, T. H. (2003). "Fruit Bats (Chiroptera: Pteropodidae) as Seed Dispersers and Pollinators in a Lowland Malaysian Rain Forest". Biotropica. 35 (4). ss. 491-502. doi:10.1111/j.1744-7429.2003.tb00606.x.
- ^ a b Popa-Lisseanu, A. G.; Delgado-Huertas, A.; Forero, M. G.; Rodríguez, A.; Arlettaz, R.; Ibáñez, C. (2007). "Bats' Conquest of a Formidable Foraging Niche: The Myriads of Nocturnally Migrating Songbirds". PLOS ONE. 2 (2). ss. e205. Bibcode:2007PLoSO...2..205P. doi:10.1371/journal.pone.0000205. PMC 1784064 $2. PMID 17299585.
- ^ Cramer, M. J.; Wilig, M. R.; Jones, C. (2001). "Trachops cirrhosus". Mammalian Species. Cilt 656. ss. 1-6. doi:10.1644/1545-1410(2001)656<0001:TC>2.0.CO;2.
- ^ Schnitzler, H.-U.; Kalko, E. K. V.; Kaipf, I.; Grinnell, A. D. (1994). "Fishing and Echolocation Behavior of the Greater Bulldog Bat, Noctilio leporinus, in the Field". Behavioral Ecology and Sociobiology. 35 (5). ss. 327-345. doi:10.1007/BF00184422.
- ^ Greenhall, A. M. (1961). Bats in Agriculture. A Ministry of Agriculture Publication. s. 8.
- ^ Wilkinson, G. (1990). "Food Sharing in Vampire Bats" (PDF). Scientific American. 262 (21). ss. 76-82. Bibcode:1990SciAm.262b..76W. doi:10.1038/scientificamerican0290-76. 21 Eylül 2020 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 16 Şubat 2020.
- ^ a b Nowak, R. M. (1991). Walker's Mammals of the World. Johns Hopkins Press. s. 1629. ISBN 978-0-8018-3970-2.
- ^ "Vampire Bats – The Good, the Bad, and the Amazing" (PDF). Natural Science Research Laboratory – Texas Tech. 15 Aralık 2017 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 14 Aralık 2017.
- ^ a b Rydell, J.; Speakman, J. R. (1995). "Evolution of nocturnality in bats: Potential competitors and predators during their early history". Biological Journal of the Linnean Society. 54 (2). ss. 183-191. doi:10.1111/j.1095-8312.1995.tb01031.x.
- ^ Esbérard, C. E. L.; Vrcibradic, D. (2007). "Snakes preying on bats: new records from Brazil and a review of recorded cases in the Neotropical Region". Revista Brasileira de Zoologia. 24 (3). ss. 848-853. doi:10.1590/S0101-81752007000300036.
- ^ Lima, S. L.; O'Keefe, J. M. (2013). "Do predators influence the behaviour of bats?". Biological Reviews. 88 (3). ss. 626-644. doi:10.1111/brv.12021. PMID 23347323.
- ^ Klimpel, S.; Mehlhorn, H. (2013). Bats (Chiroptera) as Vectors of Diseases and Parasites: Facts and Myths. Springer. s. 87. ISBN 978-3-642-39333-4. 26 Temmuz 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Şubat 2020.
- ^ a b Clayton, D. H.; Bush, S. E.; Johnson, K. P. (2015). Coevolution of Life on Hosts: Integrating Ecology and History. University of Chicago Press. s. 28. ISBN 978-0-226-30227-0. 26 Temmuz 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Şubat 2020.
- ^ "White-Nose Syndrome (WNS)". National Wildlife Health Center, U.S. Geological Survey. 30 Eylül 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2014.
- ^ Lorch, J. M.; Meteyer, C. U.; Behr, M. J.; Boyles, J. G.; Cryan, P. M.; Hicks, A. C.; Ballmann, A. E.; Coleman, J. T. H.; Redell, D. N.; Reeder, D. M.; .Blehert, D. S. (2011). "Experimental infection of bats with Geomyces destructans causes white-nose syndrome". Nature. 480 (7377). ss. 376-378. Bibcode:2011Natur.480..376L. doi:10.1038/nature10590. PMID 22031324.
- ^ "White-Nose Syndrome – Background". Canadian Cooperative Wildlife Health Centre. 16 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2014.
- ^ Daly, M. (14 Kasım 2013). "Pennsylvania's Bats Nearly Wiped Out". CBS Philadelphia. 16 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Aralık 2017.
- ^ Gutenberg, G. (7 Haziran 2012). "White-nose syndrome killing Canada's bats". Postmedia Network. 16 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 21 Nisan 2016.
- ^ "Canada : Environment Canada Announces Funding to Fight Threat of White-nose Syndrome to Bats". Mena Report. 6 Nisan 2013. 28 Mart 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2014.
- ^ "Social Bats Pay a Price: Fungal Disease, White-Nose Syndrome ... Extinction?". The National Science Foundation. 3 Temmuz 2012. 2 Ekim 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2014.
- ^ Frick, W. F.; Pollock, J. F.; Hicks, A. C.; Langwig, K. E.; Reynolds, D. S.; Turner, G. G.; Butchkoski, C. M.; Kunz, T. H. (2010). "An Emerging Disease Causes Regional Population Collapse of a Common North American Bat Species". Science. 329 (5992). ss. 679-682. Bibcode:2010Sci...329..679F. doi:10.1126/science.1188594. PMID 20689016.
- ^ "White-Nose Syndrome Confirmed in Illinois Bats: Illinois becomes 20th state in U.S. to confirm deadly disease in bats" (PDF). Illinois Department of Natural Resources. 28 Şubat 2013. 16 Şubat 2020 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 16 Şubat 2020.
- ^ "Fungus that Causes White-nose Syndrome in Bats Detected in Texas". Texas Parks and Wildlife. 23 Mart 2017. 8 Eylül 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 15 Aralık 2017.
- ^ "White Nose Syndrome of Bats Fact Sheet | Washington Department of Fish & Wildlife". wdfw.wa.gov. 3 Ocak 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Şubat 2016.
- ^ McColl, K. A.; Tordo, N.; Setien Aquilar, A. A. (2000). "Bat lyssavirus infections". Revue Scientifique et Technique. 19 (1). ss. 177-196. doi:10.20506/rst.19.1.1221. PMID 11189715.
Bats, which represent approximately 24% of all known mammalian species, frequently act as vectors of lyssaviruses
- ^ a b Calisher, C. H.; Childs, J. E.; Field, H. E.; Holmes, K. V.; Schountz, T. (2006). "Bats: Important Reservoir Hosts of Emerging Viruses". Clinical Microbiology Reviews. 19 (3). ss. 531-545. doi:10.1128/CMR.00017-06. PMC 1539106 $2. PMID 16847084.
- ^ Brüssow, H. (2012). "On Viruses, Bats and Men: A Natural History of Food-Borne Viral Infections". Viruses: Essential Agents of Life. ss. 245-267. doi:10.1007/978-94-007-4899-6_12. ISBN 978-94-007-4898-9.
- ^ a b "CDC Features – Take Caution When Bats Are Near". Centers for Disease Control and Prevention. 14 Nisan 2014. 26 Aralık 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Şubat 2020.
- ^ Eaton, Bryan T.; Broder, Christopher C.; Middleton, Deborah; Wang, Lin-Fa (2006). "Hendra and Nipah viruses: different and dangerous". Nature Reviews Microbiology. 4 (1). ss. 23-35. doi:10.1038/nrmicro1323. PMID 16357858.
- ^ Halpin, K.; Young, P. L.; Field, H. E.; Mackenzie, J. S. (2000). "Isolation of Hendra virus from pteropid bats: a natural reservoir of Hendra virus". Journal of General Virology. 81 (8). ss. 1927-1932. doi:10.1099/0022-1317-81-8-1927. PMID 10900029. 30 Ağustos 2008 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Aralık 2007.
In this paper we describe the isolation of HeV from pteropid bats, corroborating our serological and epidemiological evidence that these animals are a natural reservoir host of this virus.
- ^ Leroy, E. M.; Kumulungui, B.; Pourrut, X.; Rouque, P. (2005). "Fruit bats as reservoirs of Ebola virus". Nature. 438 (7068). ss. 575-576. Bibcode:2005Natur.438..575L. doi:10.1038/438575a. PMID 16319873.
We find evidence of asymptomatic infection by Ebola virus in three species of megabats, indicating that these animals may be acting as a reservoir for this deadly virus
- ^ Choi, C. Q. (2006). "Going to Bat". Scientific American. ss. 24, 26. 20 Kasım 2008 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Şubat 2020.
Long known as vectors for rabies, bats may be the origin of some of the most deadly emerging viruses, including SARS, Ebola, Nipah, Hendra and Marburg.
- ^ Wong, S.; Lau, S.; Woo, P.; Yuen, K.-Y. (Ekim 2006). "Bats as a continuing source of emerging infections in humans". Reviews in Medical Virology. 17 (2). John Wiley & Sons. ss. 67-91. doi:10.1002/rmv.520. PMID 17042030.
The currently known viruses that have been found in bats are reviewed and the risks of transmission to humans are highlighted.
- ^ a b Castro, J. (6 Şubat 2013). "Bats Host More Than 60 Human-Infecting Viruses". Live Science. 16 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Aralık 2017.
- ^ Dobson, A. P. (2005). "What Links Bats to Emerging Infectious Diseases?". Science. 310 (5748). ss. 628-629. doi:10.1126/science.1120872. PMID 16254175.
- ^ "Why Do Bats Transmit So Many Diseases?". IFL Science. 29 Ocak 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Aralık 2017.
- ^ Li, W.; Shi, Z.; Yu, M.; Ren, W. (28 Ekim 2005). "Bats are natural reservoirs of SARS-like coronaviruses". Science. 310 (5748). ss. 676-679. Bibcode:2005Sci...310..676L. doi:10.1126/science.1118391. PMID 16195424. Diğer özet – Science (28 Ekim 2005).
The genetic diversity of bat-derived sequences supports the notion that bats are a natural reservoir host of the SARS cluster of coronaviruses
- ^ Drosten, C.; Hu, B.; Zeng, L.-P.; Yang, X.-L.; Ge, Xing-Yi; Zhang, Wei; Li, Bei; Xie, J.-Z.; Shen, X.-R.; Zhang, Yun-Zhi; Wang, N.; Luo, D.-S.; Zheng, X.-S.; Wang, M.-N.; Daszak, P.; Wang, L.-F.; Cui, J.; Shi, Z.-L. (2017). "Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus". PLOS Pathogens. 13 (11). ss. e1006698. doi:10.1371/journal.ppat.1006698. PMC 5708621 $2. PMID 29190287.
- ^ Kerth, G.; Perony, N.; Schweitzer, F. (2011). "Bats are able to maintain long-term social relationships despite the high fission–fusion dynamics of their groups". Proceedings of the Royal Society B: Biological Sciences. 278 (1719). ss. 2761-2767. doi:10.1098/rspb.2010.2718. PMC 3145188 $2. PMID 21307051.
- ^ Fornůsková, A; Petit, E. J.; Bartonička, T.; Kaňuch, P.; Butet, A.; Řehák, Z.; Bryja, J. (2014). "Strong matrilineal structure in common pipistrelle bats (Pipistrellus pipistrellus) is associated with variability in echolocation calls". Biological Journal of the Linnean Society. 113 (4). ss. 1115-1125. doi:10.1111/bij.12381.
- ^ Carter, G. G.; Wilkinson, G. S. D. (2013). "Does food sharing in vampire bats demonstrate reciprocity?". Communicative & Integrative Biology. 6 (6). ss. e25783. doi:10.4161/cib.25783. PMC 3913674 $2. PMID 24505498.
- ^ Wilkinson, G. S. (1986). "Social Grooming in the Common Vampire Bat, Desmodus rotundus" (PDF). Anim. Behav. 34 (6). ss. 1880-1889. CiteSeerX 10.1.1.539.5104 $2. doi:10.1016/s0003-3472(86)80274-3. 22 Temmuz 2018 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 17 Şubat 2020.
- ^ a b Bohn, K. M.; Schmidt-French, Barbara; Schwartz, Christine; Smotherman, Michael; Pollak, George D. (2009). "Versatility and Stereotypy of Free-Tailed Bat Songs". PLoS ONE. 4 (8). s. e6746. Bibcode:2009PLoSO...4.6746B. doi:10.1371/journal.pone.0006746. PMC 2727915 $2. PMID 19707550.
- ^ Boughman, J. W. (1998). "Vocal learning by greater spear-nosed bats". Proceedings of the Royal Society B: Biological Sciences. 265 (1392). ss. 227-233. doi:10.1098/rspb.1998.0286. PMC 1688873 $2. PMID 9493408.
- ^ Prat, Y.; Taub, M.; Yovel, Y. (22 Aralık 2016). "Everyday bat vocalizations contain information about emitter, addressee, context, and behavior". Scientific Reports. Cilt 6. s. 39419. Bibcode:2016NatSR...639419P. doi:10.1038/srep39419. PMC 5178335 $2. PMID 28005079.
- ^ Wilkinson, G. S. (1985). "The Social Organization of the Common Vampire Bat II: Mating system, genetic structure, and relatedness" (PDF). Behavioral Ecology and Sociobiology. 17 (2). ss. 123-134. doi:10.1007/BF00299244. 13 Mart 2014 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 17 Şubat 2020.
- ^ Thomas, D. W.; Fenton, M. R.; Barclay, R. M. R. (1979). "Social Behavior of the Little Brown Bat, Myotis lucifugus: I. Mating Behavior". Behavioral Ecology and Sociobiology. 6 (2). ss. 129-136. doi:10.1007/bf00292559. JSTOR 4599268.
- ^ Keeley, A. T. H.; Keeley, B. W. (2004). "The Mating System of Tadarida brasiliensis (Chiroptera: Molossidae) in a Large Highway Bridge Colony". Journal of Mammalogy. Cilt 85. ss. 113-119. doi:10.1644/BME-004.
- ^ Toth, C. A.; Parsons, S. (2013). "Is lek breeding rare in bats?". Journal of Zoology. 291 (1). ss. 3-11. doi:10.1111/jzo.12069.
- ^ Bradbury, J. W. (1977). "Lek Mating Behavior in the Hammer-headed Bat". Zeitschrift für Tierpsychologie. 45 (3). ss. 225-255. doi:10.1111/j.1439-0310.1977.tb02120.x.
- ^ a b c Altringham 2011.
- ^ Mares, M. A.; Wilson, D. E. (1971). "Bat Reproduction during the Costa Rican Dry Season". BioScience. 21 (10). ss. 471-472+477. doi:10.2307/1295789. JSTOR 1295789.
- ^ Crichton, Elizabeth G.; Krutzsch, Philip H. (12 Haziran 2000). Reproductive Biology of Bats. Academic Press. ISBN 978-0-08-054053-5. 12 Mart 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Şubat 2020.
- ^ Neuweiler, G. (2000). Biology of Bats. Oxford University Press. s. 247. ISBN 978-0-19-509950-8.
- ^ Fenton, M. B. (1983). Just Bats. University of Toronto Press. ISBN 9781442655386.
- ^ Kunz, T. H.; Fenton, B. (2005). Bat Ecology. University of Chicago Press. s. 216. ISBN 978-0226462073.
- ^ Nagorsen, D. W.; Brigham, R. M. (1993). Bats of British Columbia. UBC Press. s. 17. ISBN 9780774804820.
- ^ Nowak, Ronald M. (1999). Walker's Mammals of the World. JHU Press. s. 269. ISBN 9780801857898.
- ^ Turbill, C.; Bieber, C.; Ruf, T. (2011). "Hibernation is associated with increased survival and the evolution of slow life histories among mammals". Proceedings of the Royal Society B. 278 (1723). ss. 3355-3363. doi:10.1098/rspb.2011.0190. PMC 3177628 $2. PMID 21450735.
- ^ a b Wilkinson, G. S.; South, J. M. (2002). "Life history, ecology and longevity in bats" (PDF). Aging Cell. 1 (2). ss. 124-131. doi:10.1046/j.1474-9728.2002.00020.x. PMID 12882342. 3 Haziran 2019 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 18 Şubat 2020.
- ^ Gager, Y.; Gimenez, O.; O'Mara, M. T.; Dechmann, D. K. N. (2016). "Group size, survival and surprisingly short lifespan in socially foraging bats". BMC Ecology. 16 (2). s. 2. doi:10.1186/s12898-016-0056-1. PMC 4714502 $2. PMID 26767616.
- ^ Eiting, T. P.; Gunnell, G. F. (2009). "Global completeness of the bat fossil record". Journal of Mammalian Evolution. 16 (3). ss. 151-173. doi:10.1007/s10914-009-9118-x.
- ^ a b c d Simmons, N. B.; Seymour, K. L.; Habersetzer, J.; Gunnell, G. F. (2008). "Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation". Nature. 451 (7180). ss. 818-821. Bibcode:2008Natur.451..818S. doi:10.1038/nature06549. hdl:2027.42/62816. PMID 18270539.
- ^ "Paleontologists Determine Original Color of Extinct Bats". SciNews. 29 Eylül 2015. 3 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 10 Eylül 2017.
- ^ Collearya, C.; Dolocanc, A.; Gardnerd, J.; Singha, Suresh; Wuttkee, M. (2015). "Chemical, experimental, and morphological evidence for diagenetically altered melanin in exceptionally preserved fossils". Proceedings of the National Academy of Sciences of the United States of America. 112 (41). ss. 12592-12597. Bibcode:2015PNAS..11212592C. doi:10.1073/pnas.1509831112. PMC 4611652 $2. PMID 26417094.
- ^ Van de Bussche, R. A.; Hoofer, S. R. (2004). "Phylogenetic relationships among recent chiropteran families and the importance of choosing appropriate out-group taxa". Journal of Mammalogy. 85 (2). ss. 321-330. doi:10.1644/1545-1542(2004)085<0321:Prarcf>2.0.Co;2.
- ^ Smith, D. "Chiroptera: Systematics". University of California Museum of Paleontology. 3 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Eylül 2017.
- ^ Eick, G. N.; Jacobs, D. S.; Matthee, C. A. (2005). "A Nuclear DNA Phylogenetic Perspective on the Evolution of Echolocation and Historical Biogeography of Extant Bats (Chiroptera)". Molecular Biology and Evolution. 22 (9). ss. 1869-1886. doi:10.1093/molbev/msi180. PMID 15930153.
- ^ Pumo, D. E. (1998). "Complete Mitochondrial Genome of a Neotropical Fruit Bat, Artibeus jamaicensis, and a New Hypothesis of the Relationships of Bats to Other Eutherian Mammals". Journal of Molecular Evolution. 47 (6). ss. 709-717. Bibcode:1998JMolE..47..709P. doi:10.1007/PL00006430. PMID 9847413.
- ^ Zhou, X. (2011). "Phylogenomic Analysis Resolves the Interordinal Relationships and Rapid Diversification of the Laurasiatherian Mammals". Systematic Biology. 61 (1). ss. 150-164. doi:10.1093/sysbio/syr089. PMC 3243735 $2. PMID 21900649.
- ^ a b c d Tsagkogeorga, G.; Parker, J.; Stupka, E.; Cotton, J. A.; Rossiter, S. J. (2013). "Phylogenomic analyses elucidate the evolutionary relationships of bats (Chiroptera)". Current Biology. 23 (22). ss. 2262-2267. doi:10.1016/j.cub.2013.09.014. PMID 24184098.
- ^ Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.; Bishop-Lilly, K. A.; Fang, X.; Wynne, J. W.; Xiong, Z.; Baker, M. L.; Zhao, W.; Tachedjian, M.; Zhu, Y.; Zhou, P.; Jiang, X.; Ng, J.; Yang, L.; Wu, L.; Xiao, J.; Feng, Y.; Chen, Y.; Sun, X.; Zhang, Y.; Marsh, G. A.; Crameri, G.; Broder, C. C.; Frey, K. G.; Wang, L.-F.; Wang, J. (2012). "Comparative Analysis of Bat Genomes Provides Insight into the Evolution of Flight and Immunity". Science. 339 (6118). ss. 456-460. Bibcode:2013Sci...339..456Z. doi:10.1126/science.1230835. PMID 23258410.
- ^ a b Teeling, E.C.; Springer, M. S.; Madsen, O.; Bates, P.; O'Brien, S. J.; Murphy, W. J. (2005). "A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record". Science. 307 (5709). ss. 580-584. Bibcode:2005Sci...307..580T. doi:10.1126/science.1105113. PMID 15681385.
- ^ Pettigrew, J. D.; Maseko, B. C.; Manger, P. R. (2008). "Primate-like retinotectal decussation in an echolocating megabat, Rousettus aegyptiacus". Neuroscience. 153 (1). ss. 226-31. doi:10.1016/j.neuroscience.2008.02.019. PMID 18367343.
- ^ Simmons, N. B.; Seymour, K. L.; Habersetzer, J.; Gunnell, G. F. (2008). "Primitive early Eocene bat from Wyoming and the evolution of flight and echolocation". Nature. 451 (7180). ss. 818-816. Bibcode:2008Natur.451..818S. doi:10.1038/nature06549. hdl:2027.42/62816. PMID 18270539.
- ^ "Bat fossil solves evolution poser". BBC News. 13 Şubat 2008. 4 Ocak 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Aralık 2017.
- ^ a b Norberg, U. M. (1994). Wainwright, P. C.; Reilly, S. M. (Ed.). Ecological Morphology: Integrative Organismal Biology. University of Chicago Press. ss. 206-208. ISBN 978-0-226-86995-7.
- ^ Bishop, K. L. (2008). "The Evolution of Flight in Bats: Narrowing the Field of Plausible Hypotheses". The Quarterly Review of Biology. 83 (2). ss. 153-169. doi:10.1086/587825. PMID 18605533.
- ^ Kaplan, Matt (2011). "Ancient bats got in a flap over food". Nature. doi:10.1038/nature.2011.9304.
- ^ Teeling, E. C.; Scally, M.; Kao, D. J.; Romagnoli, M. L.; Springer, M. S. (2000). "Molecular evidence regarding the origin of echolocation and flight in bats". Nature. 403 (6766). ss. 188-192. Bibcode:2000Natur.403..188T. doi:10.1038/35003188. PMID 10646602.
- ^ Springer, M. S.; Teeling, E. C.; Madsen, O.; Stanhope, M. J.; De Jong, W. W. (2001). "Integrated fossil and molecular data reconstruct bat echolocation". Proceedings of the National Academy of Sciences. 98 (11). ss. 6241-6246. Bibcode:2001PNAS...98.6241S. doi:10.1073/pnas.111551998. PMC 33452 $2. PMID 11353869.
- ^ L., G.; Wang, J.; Rossiter, S. J.; Jones, G.; Zhang, S. (2007). "Accelerated FoxP2 evolution in echolocating bats". PLOS ONE. 2 (19). ss. e900. Bibcode:2007PLoSO...2..900L. doi:10.1371/journal.pone.0000900. PMC 1976393 $2. PMID 17878935.
- ^ DesRoche, K.; Fenton, M. B.; Lancaster, W. C. (2007). "Echolocation and the thoracic skeletons of bats: a comparative morphological study". Acta Chiropterologica. 9 (2). ss. 483-494. doi:10.3161/1733-5329(2007)9[483:EATTSO]2.0.CO;2.
- ^ Li, G.; Wang, J.; Rossiter, S. J.; Jones, G.; Cotton, J. A.; Zhang, S. (2008). "The hearing gene Prestin reunites the echolocating bats". Proceedings of the National Academy of Sciences of the United States of America. 105 (37). ss. 13959-13964. Bibcode:2008PNAS..10513959L. doi:10.1073/pnas.0802097105. PMC 2544561 $2. PMID 18776049.
- ^ Wang, Zhe; Zhu, Tengteng; Xue, Huiling; Fang, Na; Zhang, Junpeng; Zhang, Libiao; Pang, Jian; Teeling, Emma C.; Zhang, Shuyi (2017). "Prenatal development supports a single origin of laryngeal echolocation in bats". Nature Ecology & Evolution. 1 (2). s. 21. doi:10.1038/s41559-016-0021. PMID 28812602.
- ^ Lei, M.; Dong, D. (2016). "Phylogenomic analyses of bat subordinal relationships based on transcriptome data". Scientific Reports. Cilt 6. s. 27726. Bibcode:2016NatSR...627726L. doi:10.1038/srep27726. PMC 4904216 $2. PMID 27291671.
- ^ Neuweiler, Gerhard (2000). "Phylogeny and systematics". The Biology of Bats. Oxford University Press. ss. 287-299. ISBN 978-0195099508.
- ^ a b c Prothero, D. R. (2017). "Laurasiatheria: Chiroptera". The Princeton Field Guide to Prehistoric Mammals. Princeton University Press. ss. 112-116. ISBN 978-0-691-15682-8.
- ^ Hutcheon, J. M.; Garland, T. (2004). "Are Megabats Big?". Journal of Mammalian Evolution. 11 (3/4). s. 257. doi:10.1023/B:JOMM.0000047340.25620.89.
- ^ Holland, R. A. (2004). "Echolocation signal structure in the Megachiropteran bat Rousettus aegyptiacus Geoffroy 1810". Journal of Experimental Biology. 207 (25). ss. 4361-4369. doi:10.1242/jeb.01288. PMID 15557022.
- ^ Brown, W. M. (2001). "Natural selection of mammalian brain components". Trends in Ecology and Evolution. 16 (9). ss. 471-473. doi:10.1016/S0169-5347(01)02246-7.
- ^ a b Stephen, J.; Olney, P. (1994). Creative Conservation: Interactive Management of Wild and Captive Animals. Springer. ss. 352. ISBN 978-0412495700.
- ^ Wilson, D.E.; Mittermeier, R.A., (Ed.) (2019). Handbook of the Mammals of the World – Volume 9. Barselona: Lynx Ediciones. ss. 1-1008. ISBN 978-84-16728-19-0.
- ^ "Mission & Vision". Bat Conservation International. 4 Ağustos 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Kasım 2017.
- ^ "Bats and the Law". Bat Conservation Trust. 31 Ekim 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Kasım 2017.
- ^ a b "Wildlife Protection Ordinance 1998" (PDF). FAO. 17 Kasım 2017 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 16 Kasım 2017.
- ^ Leong, T. M.; Teo, S. C.; Lim, K. K. P. (2009). "The Naked Bulldog Bat, Cheiromeles torquatus in Singapore — past and present records, with highlights on its unique morphology (Microchiroptera: Molossidae)". Nature in Singapore. Cilt 2. ss. 215-230.
- ^ Ceballos, G.; Ehrlich, A. H.; Ehrlich, P. R. (2015). The Annihilation of Nature: Human Extinction of Birds and Mammals. Johns Hopkins University Press. ss. 75-76. ISBN 978-1421417189.
- ^ "All about bats". Bat Conservation International. 24 Ocak 2002. 23 Haziran 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Şubat 2020.
- ^ "Welcome to the World's Largest Occupied Bat Houses". Florida Museum of Natural History. 30 Ocak 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Aralık 2017.
- ^ "Protecting and managing underground sites for bats, see section 6.4" (PDF). Eurobats. 12 Mayıs 2012 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 18 Mayıs 2006.
- ^ "Pillbox converted to bat retreat". BBC. 6 Nisan 2006. 15 Ocak 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 18 Mayıs 2006.
- ^ "Bypass wings it with bat bridges". BBC. 4 Nisan 2008. 19 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 21 Ağustos 2016.
- ^ "Bat bridges cost £27k per animal". BBC. 22 Ekim 2009. 19 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 21 Ağustos 2016.
- ^ "Agency Guide to Cave and Mine Gates 2009" (PDF). Batcon.org. 20 Ekim 2012 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 1 Kasım 2017.
- ^ Pfeiffer, Martin J. (Şubat 2019). Bats, People, and Buildings: Issues and Opportunities. Madison, WI: United States Department of Agriculture, Forest Service, Forest Products Laboratory. Erişim tarihi: 26 Mart 2019.
- ^ Hopkins, J.; Bourdain, A. (2004). Extreme Cuisine: The Weird & Wonderful Foods that People Eat. Periplus. s. 51. ISBN 978-0-7946-0255-0. 17 Şubat 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Şubat 2020.
- ^ Baerwald, E. F.; D'Amours, G. H.; Klug, B. J.; Barclay, R. M. R. (2008). "Barotrauma is a significant cause of bat fatalities at wind turbines". Current Biology. 18 (16). s. R695–R696. doi:10.1016/j.cub.2008.06.029. PMID 18727900.
- ^ a b "B.C. study to help bats survive wind farms". National Wind Watch. 23 Eylül 2008. 19 Nisan 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Nisan 2015.
- ^ "Bats take a battering at wind farms 10 Kasım 2013 tarihinde Wayback Machine sitesinde arşivlendi.", New Scientist, 12 Mayıs 2007
- ^ "Caution Regarding Placement of Wind Turbines on Wooded Ridge Tops" (PDF). Bat Conservation International. 4 Ocak 2005. 23 Mayıs 2006 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 21 Nisan 2006.
- ^ Arnett, E. B.; Erickson, W. P.; Kerns, J.; Horn, J. (12 Haziran 2005). "Relationships between Bats and Wind Turbines in Pennsylvania and West Virginia: An Assessment of Fatality Search Protocols, Patterns of Fatality, and Behavioral Interactions with Wind Turbines" (PDF). Bat Conservation International. 10 Şubat 2006 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 21 Nisan 2006.
- ^ Baerwald, E. F.; D'Amours, G.H.; Klug, Brandon J.; Barclay, R. M. R. (26 Ağustos 2008). "Barotrauma is a significant cause of bat fatalities at wind turbines". Current Biology. 18 (16). ss. R695-R696. doi:10.1016/j.cub.2008.06.029. OCLC 252616082. PMID 18727900.
- ^ Johnson, J. B.; Ford, W. M.; Rodrigue, J. L.; Edwards, J. W. (2012). Effects of Acoustic Deterrents on Foraging Bats. U.S. Department of Agriculture, U.S. Forest Service, Northern Research Station. ss. 1-5.
- ^ McCracken, G. F. (1993). "Folklore and the Origin of Bats". BATS Magazine. Bats in Folklore. 11 (4).
- ^ Chwalkowski, F. (2016). Symbols in Arts, Religion and Culture: The Soul of Nature. Cambridge Scholars Publishing. s. 523. ISBN 978-1443857284.
- ^ Saleh, A. (19 Temmuz 2001). "Sex-mad 'ghost' scares Zanzibaris". BBC News. 13 Mart 2012 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Aralık 2014.
- ^ "Aztec Symbols". Aztec-history.net. 15 Mart 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Haziran 2013.
- ^ Read, K. A.; Gonzalez, J. J. (2000). Mesoamerican Mythology. Oxford University Press. ss. 132-134. ISBN 978-0195149098.
- ^ "Artists Inspired by Oaxaca Folklore Myths and Legends". Oaxacanwoodcarving.com. 10 Kasım 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 June 2013.
- ^ Arnott, K. (1962). African Myths and Legends. Oxford University Press. ss. 150-152.
- ^ "Chinese symbols" (PDF). British Museum. 24 Mayıs 2019 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 10 Eylül 2017.
- ^ Grant, G. S. "Kingdom of Tonga: Safe Haven for Flying Foxes". Batcon.org. 12 Ağustos 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Haziran 2013.
- ^ Cartwright, M. (28 Ekim 2013). "Zapotec Civilization". Ancient History Encyclopedia. 21 Ocak 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Aralık 2017.
- ^ de Vries, A. (1976). Dictionary of Symbols and Imagery. Amsterdam: North-Holland. s. 36. ISBN 978-0-7204-8021-4.
- ^ Miller, Elizabeth (1998). "Bats, Vampires & Dracula". Newsletter of the Florida Bat Conservation Centre, Sonbahar 1998. 24 Ocak 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Aralık 2017.
- ^ Fleisher, M. L. (1976). The Encyclopedia of Comic Book Heroes Volume 1 Batman. Collier Books. s. 31. ISBN 978-0-02-080090-3.
- ^ Tramoyeres Blasco, L. "Lo Rat Penat en el escudo de armas de Valencia". 14 Mart 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Kasım 2014.
- ^ Alomar i Canyelles, A. I. (1998). L'Estendard, la festa nacional més antiga d'Europa. Palma. ss. XIII-XXI.
- ^ "Estudio de los escudos recogidos en la orla de latabla de la magistratura" (PDF). Emblemata Revista Aragonesa de Emblematica. Cilt 6. 2005. s. 242. 10 Eylül 2017 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 19 Şubat 2020.
- ^ "Official state bats". Netstate. 9 Mart 2011 tarihinde kaynağından arşivlendi. Erişim tarihi: 13 Şubat 2011.
- ^ Boyles, Justin G.; Cryan, Paul M.; McCracken, Gary F.; Kunz, Thomas H. (2011). "Economic Importance of Bats in Agriculture". Science. 332 (6025). ss. 41-42. Bibcode:2011Sci...332...41B. doi:10.1126/science.1201366. PMID 21454775.
- ^ Weaver, H. D. (2008). Missouri Caves in History and Legend. University of Missouri Press. ss. 64-69. ISBN 978-0-8262-6645-3.
- ^ Whisonant, R. C. (2001). "Geology and History of Confederate Saltpeter Cave Operations in Western Virginia" (PDF). Virginia Minerals. 47 (4). ss. 33-43. 27 Aralık 2016 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 19 Şubat 2020.
- ^ Christensen, RaeAnn. "Best time to see the bat colony emerge from Congress Bridge in Downtown Austin". Fox7. 19 Ekim 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 21 Ağustos 2016.
Genel
- Altringham, J. D. (2011). Bats: From Evolution to Conservation. Oxford University Press. ISBN 978-0199207114.
- Fenton, M. B. (2001). Bats. Checkmark Books. ISBN 978-0-8160-4358-3.
- Fenton, M. B.; Simmons, N. B. (2015). Bats: A World of Science and Mystery. University of Chicago Press. ISBN 978-0226065120.
Dış bağlantılar
- UK Bat Conservation Trust15 Ekim 2018 tarihinde Wayback Machine sitesinde arşivlendi.
- Tree of Life26 Temmuz 2020 tarihinde Wayback Machine sitesinde arşivlendi.
- Microbat Vision29 Ocak 2020 tarihinde Wayback Machine sitesinde arşivlendi.
- The DSP Behind Bat Echolocation28 Temmuz 2020 tarihinde Wayback Machine sitesinde arşivlendi. – Çeşitli yarasa ekolokasyonlarının analizi















