Transkripsiyon (genetik)
Transkripsiyon, yazılma veya yazılım, DNA'yı oluşturan nükleotit dizisinin RNA polimeraz enzimi tarafından bir RNA dizisi olarak kopyalanması sürecidir. Başka bir deyişle, DNA'dan RNA'ya genetik bilginin aktarımıdır. Protein kodlayan DNA durumunda, transkripsiyon, DNA'da bulunan genetik bilginin (bir mesajcı RNA aracılığıyla) bir protein veya peptit dizisine çevirisinin ilk aşamasıdır. RNA'ya yazılan bir DNA parçasına "transkripsiyon birimi" denir. Transkripsiyonda hata kontrol mekanizmaları vardır, ama bunlar DNA çoğalmasındakinden daha az sayıda ve etkindirler; dolayısıyla transkripsiyon DNA çoğalması kadar aslına sadık değildir.
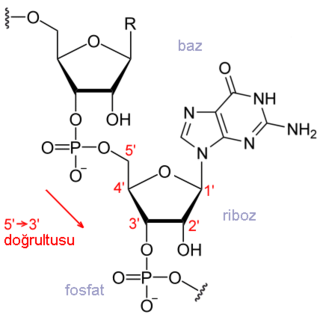
Başlama noktasından önceki diziler upstream,sonraki diziler downstream olarak adlandırılır.
DNA sentezinde olduğu gibi transkripsiyonda da RNA sentezi 5' → 3'doğrultusunda ilerler. Yani,upstream'dan downstream'e doğru ilerler
DNA'da bulunan bilgi işlevsel protein veya RNA ürünlerinin sentezini sağlar. Bu işlevsel ürünleri kodlayan DNA dizilerine gen, bunların oluşumuna da "gen ifadesi" denir. DNA'daki bilginin RNA olarak yazılmış haline "transkript" denir. Ribozomların protein sentezi yapmak için okuduğu RNA molekülü "mesajcı RNA"dır. Prokaryotlarda RNA polimerazın ürettiği RNA ile ribozomların okuduğu mRNA aynı moleküldür. Ökaryotlarda ise transkript bir takım işlemlerden geçtikten sonra olgun mRNA olur. Bu bakımdan, işlem görmemiş mRNA'ya "öncül mRNA", "prekürsör mRNA" veya "pre-mRNA" da denir.
Aşağıda ökaryotik ve prokaryotik organizmalardaki transkripsiyonun benzer ve farklı yönleri ele alınarak konuya genel bir bakış verilmektedir. Arkelerin transkripsiyon mekanizması ökaryotlarınkine benzer. Ayrıntılar için prokaryotik transkripsiyon ve ökaryotik transkripsiyon maddelerine bakınız.
Gen ifadesinin düzenlenmesi
Bir genin okunmaya başlandığı noktanın hemen yukarısındaki bölgenin adı "promotör"dür ("Yukarı" ve "aşağı" terimleri transkripsiyon yönüne bağlı olarak kullanılır: transkripsiyon yönü aşağıdır, transkripsiyon yönünün tersi yukarıdır.). Promotör bölgesinde genlerin ifadesini kontrol eden DNA dizileri yer alır. Ökaryotlarda promotör bölgelerden başka, "hızlandırıcı" (İngilizce enhancer) adı verilen DNA bölgeleri de gen ifadesine etki eder. Bu hızlandırıcılar transkripsiyon başlama noktasından çok uzakta olsalar da üç boyutlu uzayda ona yakındırlar. Promotörlere ve hızlandırıcılara bağlanan bazı transkripsiyon faktörleri RNA polimerazla etkileşerek onun çalışmasını engeller veya onu uyarırlar.
RNA polimeraz
Ökaryotik transkripsiyonda üç farklı RNA polimeraz vardır, bunlar farklı sınıf genleri okumaktan sorumludur.
| RNA Polimeraz I | 45S ribosomal RNA (rRNA) genleri |
| RNA polimeraz II | Mesajcı RNA (mRNA) genleri |
| RNA Polimeraz III | Taşıyıcı RNA (tRNA), 5S rRNA ve bazı başka küçük RNA genleri |
Prokaryotik transkripsiyonda bütün genler tek bir RNA polimeraz tarafından okunur. Arkelerin de bir RNA polimerazı vardır ama çalışma mekanizması ökaryotik RNA polimerazlarınki gibidir. Çok alt birimli olan bu RNA polimerazların yanı sıra SP67 ve T7 gibi fajların ve mitokondrilerin kendilerine has, tek alt birimli RNA polimerazları vardır.
Prokaryot polimerazı dört alt birimden (α2, β, β' ve ω) oluşur. "Sigma (σ)" olarak adlandırılan bir diğer protein ise RNA polimerazın belli promotörlere bağlanmasını sağlar ama RNA'nın sentezi için gerekli değildir. Sigmanın birkaç çeşidi vardır ve hangi genin okunacağı RNA polimeraza bağlı olan sigma alt biriminin türüne bağlıdır. Ökaryotik polimerazların daha fazla sayıda alt birimi vardır.[1]
Bir prokaryot olan E. coli'nin RNA polimerazı en çok ökaryot RNA polimeraz II'ye benzer ve bunların evrimsel olarak ortak bir geçmişe sahip oldukları muhtemel görülür.
RNA polimeraza yardımcı olan çeşitli kofaktör proteinler vardır. Tüm promotörlerden yapılan yazılmada rol oynayan bu proteinlere genel transkripsiyon faktörleri denir. Bunların hata kontrolü veya DNA tamiri gibi yardımcı işlevleri vardır. Diğer kofaktörler RNA polimerazın belli bazı genleri ifade edip etmeyeceğini belirler; bunlara sadece transkripsiyon faktörü denir. Gen ifadesini engelleyici transkripsiyon faktörlerine "represör", kolaylaştırıcı olanlara "aktivatör" denir. Bu sayede bir genin farklı metabolik şartlarda veya farklı dokularda uygun düzeyde ifadelenmesi mümkün olur.
Mekanizma
Prokaryot ve ökaryotlarda transkripsiyon mekanizmalarının ayrıntıları farklılık gösterir. Prokaryotların çekirdek zarları olmadığı için, oluşmakta olan RNA'nın aynı anda ribozomlar tarafından da okunup çevrimi yapılabilir. Oysa ökaryotlarda, RNA çekirdek içinde oluştuktan sonra ribozomların bulunduğu sitoplazma ve endoplazmik retikuluma taşınır. Dolayısıyla transkripsiyon ve translasyon farklı mekân ve zamanlarda gerçekleşir.
Transkripsiyon üç aşamadan oluşur: başlama, uzama ve sonlanma. Buna ek olarak ökaryotlarda bir işlenme aşaması vardır.
Başlama
Sitoplazmada translasyonun başlaması için gerekli genetik şifre mRna aracılığı ile çekirdek zarından geçerek sitoplazmaya varır. Başlangıç kodonu olarak adlandırılan (AUG) dizilimi ile tRna lar aminoasit taşımına başlar. Başlangıç kodonu ile metiyonin aminoasidi eşleşir.
Prokaryotlar: Prokaryot promotörlerinde iki önemli DNA dizisi vardır: biri, transkripsiyon başlama noktasından 10 nükleotit yukarıda (-10 konumunda) olan TATAAT dizisi; öbürü de -35'te bulunan TTGACA dizisi. Prokaryotlarda RNA polimeraz DNA'ya bağlanır, sonra bir promotör bulana kadar onun üzerinde ilerler. Sigma altbirimi -35 dizisini tanıyıp RNA polimerazın daha sıkı bağlanmasını sağlar. Sonra sigma ayrılır ve geriye dört alt birimli çekirdek enzimi birakır. A-T baz çiftleri G-C baz çiftlerine kıyasla daha zayıf oldukları için -10 dizisinde DNA zincirleri birbirlerinden ayrılırlar. İki DNA zincirinin birbirinden ayrıldığı bölge "transkripsiyon kabarcığı" olarak tabir edilir. RNA polimeraz uygun noktadan itibaren RNA sentezine başlar.
Ökaryotlar: Ökaryotlarda -30'da TATAAA veya benzeri bir dizi (TATA kutusu) ve -80 civarında bulunan GGCCAATCT dizisi (CCAAT kutusu) vardır.
Ökaryotlardaki TATA kutusuna önce TATA Bağlanma Proteini (TBP) bağlanır. Bu başlama kompleksi RNA polimerazı promotöre seferber eder ve oradan transkripsiyon sürecini başlatmasını sağlar. Bu proteinler temel düzeyde bir transkripsiyon için yeterlidirler. Daha yüksek seviyede transkripsiyon elde etmek için başka transkripsiyon faktörleri gereklidir.
Promotör ve ökaryotlarda hızlandırıcılara bağlanan düzenleyici proteinler, RNA polimerazın DNA'ya bağlanmasına engel olarak veya bağlanmasını kolaylaştırarak transkripsiyonun seviyesini düzenlerler.
Uzama
Uzama, prokaryot ve ökaryotlarda benzer şekilde gerçekleşir. Uzayan RNA zincirinin 3' ucuna nükleotitler eklenir. Yani, gelen nükleotidin 5' fosfat grubu ile RNA zincirindeki 3' hidroksil grubu arasında bir fosfodiester bağı oluşur. İki DNA zincirinden sadece biri, kendisini tümleyici bir RNA ipliğinin sentezi için kullanılır; buna "şablon zincir" denir. Sentez sırasında geçici bir DNA-RNA ikilisi oluşur ama sonra RNA DNA'dan ayrışır ve ilerleyen enzimin gerisinden DNA tekrar kapanıp normal çift sarmallı haline geri döner.
Sonlanma

Prokaryotlar: RNA polimeraz bir sonlanma sinyaline rastlayınca RNA sentezi sona erer. Prokaryotik genlerde iki tip sonlanma vardır: "ro" adı verilen sonlanma proteininin gerekli olup olmadığına göre, ro'ya bağlı ve ro'dan bağımsız sonlanma. Bunların sinyalleri farklıdır. Ro'dan bağımsız sonlanmada sık G/C nükleotitli bir bölgeyi izleyen sık A/T'li bir bölge bulunur. G/C'li kısım RNA'ya yazılınca, oradaki nükleotitler firkete görünümlü bir şekil alırlar ve bu RNA polimerazı yavaşlatır. Bunu izleyen sık A/T'li kısımda ise polimeraz duraklar ve DNA'dan kopar.
Ro'ya bağlı sonlanmada ise DNA'da sık C'li bir bölge olur. Transkripsiyon sırasında ro proteini büyümekte olan RNA'ya bağlanıp, onun üzerinden polimeraza doğru ilerlemeye başlar. Polimeraz sık C'li bölgeye gelince duraklar, bu sayede ro polimeraza yetişir ve yeni sentezlenmiş RNA'yı ondan kopartır.
Ökaryotlar: Ökaryotlarda prokaryotlardaki gibi belirgin sonlanma sinyalleri yoktur. RNA polimeraz mRNA'nın biteceği yerden 1000-2000 nükleotit daha ileriye kadar okumaya devam eder. Bu RNA sonradan işlenerek fazla uzamış kısmı çıkartılır.
İşlenme
Prokaryot RNAlar sentezlendikten sonra herhangi bir işlemden geçmeden ribozomlar tarafından okunarak protein sentezinde kullanılırlar; hatta bir RNA'nın sentezi bitmeden bir ribozom onun çevirisini yapmaya başlar.
Ökaryotlarda en son mRNA'nın oluşması için sınıf II RNA polimeraz okumaları (transkriptleri) bir takım işlemlerden geçer. Bu işlemler arasında başlık takılması (İngilizce capping), poliadenilasyon ve intron çıkarılması (uç birleştirme; İngilizce splicing) vardır. Ribozomal ve taşıyıcı RNAlar da işlenir, ama ne başlık alırlar ne de poliadenile olurlar.
Başlık RNA'nın 5' ucunda olur. RNA'ya 5'-5' fosfodiester bağlantısı ile metilli bir guanin nükleotidi eklenir. Bu "başlık", mRNA'nın çeviri sırasında ribozomlar tarafından tanınması için önemlidir.
Poliadenilasyonda RNA'nın ucu kesilerek doğru olan 3' uç ortaya çıkar ve buna bir dizi adenin nükleotiti eklenir. 3' ucun konumu RNA içinde bulunan bir nükleotit dizisi tarafından belirlenir. Bu dizi, AAUAAA, poliadenilasyon sinyali olarak adlandırılır. Gerekli enzimler bu diziyi tanıyınca RNA bu sinyalden 10 - 30 nükleotit aşağıda kesilir ve sonra ona bir dizi adenin eklenir. Bu adeninlerin eklenmesinde bir şablon kullanılmaz; A'lar sadece peşpeşe RNA'nın 3' ucuna eklenir. Bu poli(A) kuyruğu ortalama 200 nükleotit uzunluğunda olur ve RNA'yı yıkımdan korur.
İntronlar, uçbirleştirme (ing. splicing) işlemi sonucu prekürsör RNA'dan çıkartılan bölümlerdir, kalan kısımlar ekson olarak adlandırılır. Çıkartılma mekanizmasına bağlı olarak iki tip introndan söz edilir. Tip I intronlarda RNA'nın katalizör özelliği vardır; kendi kendini kesip birleştirme yeteneğine sahiptir. Tip II intronlarda bu işlemden sorumlu olan splisozom (İngilizce spliceosome) adlı büyük bir RNA/protein kompleksi vardır. Splisozom, intron-ekson sınırını tanıyıp RNA'yı o noktada keser, sonra da bitişik eksonları birleştirerek ergin mRNA'yı meydana getirir.
Ters transkripsiyon
Bazı virüsler (örneğin AIDS hastalığına neden olan HIV) RNA'yı DNA'ya yazar. Bu tür yazılma ters transkriptaz adlı enzim tarafından gerçekleştirilir. HIV'da ters transkriptaz, viral genomdan bir tümleyici (komplementer) DNA ipliği (cDNA) sentezler. Başka bir enzim, ribonükleaz H, RNA ipliğini sindirir. Ardından ters transkriptaz, cDNA'yı tümleyici bir DNA ipliği daha sentezleyerek çift sarmallı bir DNA oluşturur. Bu viral DNA, entegraz adlı bir enzim aracılığıyla konak hücrenin genomuna dahil olur. Bu sürecin sonucunda konak hücre yeni virüslerin oluşumu için gerekli olan viral proteinleri ve RNA ipliğini üretmeye başlar. Ardından hücre programlanmış ölüm mekanizmasıyla (apoptoz) imha olur.
Tarihçe
RNA polimerazın in vitro olarak RNA sentezlediği çeşitli laboratuvarlarda 1965'te gösterilmiştir. Roger Kornberg ökaryotik transkripsiyon mekanizmasının moleküler ayrıntıları üzerinde yaptığı çalışmalardan dolayı Nobel Kimya Ödülü'nü kazanmıştır.
Notlar
- ^ Dirk Eick, Andrew Wedel and Hermann Heumann (1994). "From initiation to elongation: comparison of transcription by prokaryotic and eukaryotic RNA polymerases". Trends in Genetics. 10 (8). ss. 292-296.
Kaynakça
- Molecular Cell Biology. 4th ed. Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James E. New York: W. H. Freeman & Co.; c2000.
- Molecular Biology of the Cell 4th ed. Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter. New York and London: Garland Science; c2002
- The Cell - A Molecular Approach. 2nd ed. Cooper, Geoffrey M. Sunderland (MA): Sinauer Associates, Inc; c2000.
- Lehninger Principles of Biochemistry, 4th edition, David L. Nelson & Michael M. Cox
- Principles of Nuclear Structure and Function, Peter R. Cook
- Essential Genetics, Peter J. Russell