Struthiomimus
| Struthiomimus | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Montelenmiş iskeleti, Oxford Üniversitesi Doğa Tarihi Müzesi | |||||||||||||||||||
| Biyolojik sınıflandırma | |||||||||||||||||||
| |||||||||||||||||||
| İkili adlandırma | |||||||||||||||||||
| †Struthiomimus altus Osborn, 1916 | |||||||||||||||||||
Struthiomimus (anlamı: "Devekuşu taklitçisi", Antik Yunanca kökenli olan ve "devekuşu" anlamına gelen στρούθειος / stroutheios ve "mimik" veya "taklitçi" anlamına gelen μῖμος / mimos sözcüklerinden oluşur), Geç Kretase döneminde Kuzey Amerika'da yaşamış ornithomimid ailesinden bir dinozor cinsi. Ornithomimidler, dişsiz bir gagaya sahip, uzun bacaklı ve iki ayaklı canlılardı, özetle devekuşu benzeri dinozorlardı. Tip türü, Struthiomimus altus , Dinozor Eyalet Parkı'nda bulunan yaygın küçük dinozorlardan biridir; ilgili fosil alanında sayılarının fazlalığı bu hayvanların saf etoburlardan ziyade otobur veya hepçil olduklarını düşündürür.[1]
Keşif

,
1901'de Lawrence Lambe daha sonra holotip CMN 930 olarak tanımlanacak bazı tam olmayan kalıntılar buldu ve onları Ornithomimus altus olarak adlandırıp onları daha önce 1890'da Othniel Charles Marsh tarafından tarif edilen kalıntılarla aynı cinse yerleştirdi. Bu dinozorun tür adı, "yüce" veya "asil" anlamına gelen Latince altus sözcüğüdür. Bununla birlikte, 1914'te, Alberta'daki Kızıl Geyik Irmağı sahasında Barnum Brown tarafından neredeyse tam bir iskelet (AMNH 5339) keşfedildi ve bu, O. altus'un Henry Fairfield Osborn tarafından yeni bir alt cins olan Struthiomimus'un tip cinsi olarak tanımlanmasına neden oldu. 1917.[2] Dale Russell, 1972'de Struthiomimus'u tam bir cins yaptı ve aynı zamanda diğer birkaç numuneyi de ona atfetti: AMNH 5375, AMNH 5385, AMNH 5421, CMN 8897, CMN 8902 ve ROM 1790, tüm kısmi iskeletler.[3] 1916'da Osborn ayrıca Ornithomimus tenuis (Marsh, 1890) Struthiomimus tenuis olarak yeniden adlandırdı.[2] Bu durum bugün bir nomen dubium olarak kabul edilmektedir. 2016 yılında, ROM 1790 adlı fosil numune yeni bir cins ve türün holotipi haline getirildi (Rativates evadens).[4]

Sonraki yıllarda William Arthur Parks, Struthiomimus'un diğer dört türünü adlandırdı: Struthiomimus brevetertius Parks 1926,[5] Struthiomimus samueli Parks 1928,[6] Struthiomimus currellii Parks 1933 ve Struthiomimus ingens Parks 1933.[7] Bunlar bugün ya Dromiceiomimus'a ya da Ornithomimus'a ait olarak görülmektedir.
1997'de Donald Glut, Struthiomimus lonzeensis adından bahsetmiştir.[8] Bu muhtemelen bir lapsus calami yani Ornithomimus lonzeensis için bir hataydı (Dollo 1903) Kuhn 1965. Struthiomimus altus, Geç Kampaniyen yaşlı (Judith çağı) Oldman Formasyonu'ndan gelir.[9]
Olası bir ikinci Struthiomimus türü, erken Maastrihtiyen (Edmontoniyen yaşlı) Horseshoe Kanyon Formasyonu'ndan bilinmektedir. Dinozor faunasının hızlı bir dönüşüm gösterdiği düşünüldüğünde, onlara yeni bir ad verilmemiş olmasına rağmen daha genç Struthiomimus örneklerinin S. altus'tan ayrı bir türü temsil etmesi olası görünmektedir.[9][10]
Alt Lance Formasyonundan ek Struthiomimus numuneleri ve eşlenikleri (boyutça Gallimimus'a benzer olanlar) Ornithomimus'takilere benzer olarak daha düz ve daha uzun el pençelerine sahip olma eğilimindedir. Nispeten eksiksiz bir Lance Formasyonu örneği olan BHI 1266 numunesi, orijinal olarak Ornithomimus sedens'e (1892'de Marsh tarafından adlandırılan[11] ) atfedilmiştir ve daha sonra Struthiomimus sedens olarak sınıflandırılmıştır.[12] Van der Reest ve arkadaşlarının 2015 tarihli bir makalesinde BHI 1266'yı Ornithomimus sp. olarak listelenirken,[13] aynı yıl başka bir makalede ise Struthiomimus sp. olarak düşünülmüştür. Her iki cinsin yeniden değerlendirilmesi beklenmektedir.[9]
Fiziksel Özellikler
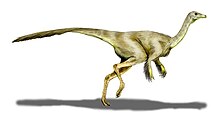
S. altus'un vücut uzunluğunun yaklaşık 4,3 metre olduğu ve yüksekliğinin kalça hizasında 1,4 m kadar olduğu tahmin edilmektedir. Bunun yanında ağırlığının ise yaklaşık 150 kg geldiği düşünülmektedir.[14] S. sedens''in boyunun ise yaklaşık 5,8 m uzunlukta, kalçadan yüksekliğininse 1,95 m kadar geldiği tahmin edilmiştir ve vücut ağırlığı da hemen 420 kg ile onu Kuzey Amerika'nın en büyük ornitomimmid türü yapmaktadır.[15] Struthiomimus, ornithomimidlere özgü bir fizik ve iskelet yapısına sahiptir ve bu noktada vücut orantısı ve anatomik ayrıntılarla Ornithomimus ve Gallimimus gibi yakından ilişkili cinslerden farklılaşmaktadır.[16]

Diğer ornithomimidlerde olduğu gibi, uzun boyunlarında küçük ince kafaları vardı (kalçalarının önünden itibaren vücut uzunluğunun yaklaşık %40'ını oluşturmaktadır.).[17] Gözleri büyüktü ve çeneleri dişsizdi. Onların vertebra sütunları, on boyun omuru, on üç sırt omuru, altı kalça omuru ve yaklaşık otuz beş kuyruk omurundan oluşuyordu.[18] Kuyrukları nispeten sertti ve muhtemelen denge için kullanılıyordu.[2] Struthiomimus, narin kol ve ellere sahiptir. Kol kemikleri hareketsizdir ve ilk parmağıyla diğer parmakları arasında sınırlı şekilde karşıtlık vardır.[19] Diğer ornithomimidlerde olduğu gibi, ancak teropodlar arasında alışılmadık bir şekilde, üç parmak kabaca aynı uzunluktaydı ve pençeler sadece hafifçe kavisliydi; Henry Fairfield Osborn , 1917'de bir S. altus iskeletini betimleyerek, onların kol yapısını bir tembel hayvanınkiyle karşılaştırdı.[2] Bunların kanat tüylerini desteklemek için yapılan evrimsel uyarlamalar olduğu düşünülmektedir.[20] Struthiomimus, yakın akrabalarından yalnızca anatomisinin ince yönlerinde farklıydı. Üst gaganın kenarı, düz gaga kenarları olan Ornithomimus'un aksine , Struthiomimus'ta içbükeydir.[10] Struthiomimus, diğer ornithomimidlere göre humerus kemiğine oranla daha uzun ellere sahipti.[17] Ön ayakları Ornithomimus'unkilerden daha sağlamdı.[10]
Sınıflandırma
Struthiomimus; Anserimimus, Archaeornithomimus, Dromiceiomimus, Gallimimus, Ornithomimus ve Sinornithomimus'u da içeren bir grup olan Ornithomimidae ailesinin bir üyesidir.
Struthiomimus'un fosil kalıntıları yanlışlıkla Ornithomimus'a atandığında, Struthiomimus'un ait olduğu daha büyük grup olan Ornithomimosauria da yıllar içinde birçok değişikliğe uğramıştır. Örneğin, O.C. Marsh başlangıçta Struthiomimus'u, teropodlarla yakından ilişkili olmayan büyük bir dinozor klanı olan Ornithopoda'ya dahil etmiştir.[21] Beş yıl sonra Marsh, Struthiomimus'u Ceratosauria'da sınıflandırdı.[22][23] 1891'de Baur, bu cinsi Iguanodontia içine yerleştirdi.[24] 1993 gibi geç bir tarihte, Struthiomimus, Oviraptorosauria'ya atfedilmiştir.[25] Bununla birlikte, 1990'larda, Struthiomimus'u Coelurosauria içine yerleştiren çok sayıda çalışma vardı.[26][27][28][29]
Ornithomimidler ve diğer teropodlar arasındaki farkı tanıyarak Rinchen Barsbold, 1976'da ornithomimidleri kendi alt takımlarına (Ornithomimosauria) koymuştur.[30] Ornithomimidae ve Ornithomimosauria'nın seçim bölgesi farklı yazarlara göre değişiyordu. Örneğin Paul Sereno, Ornithomimidae'yi 1998'de tüm ornithomimosaurları dahil etmek için kullandı, ancak daha sonra Ornithomimosauria içinde daha özel bir tanıma (gelişmiş ornithomimosaurlar) geçildi,[31] bu yüzyılın başında diğer yazarlar tarafından bir sınıflandırma şeması benimsenmiştir.
Xu ve arkadaşlarının 2011'deki analizini takip eden kladogram:[32]
| Ornithomimidae |
| ||||||||||||||||||||||||||||||||||||
Paleobiyoloji
Bruce Rothschild ve diğer paleontologlar tarafından 2001 yılında yürütülen bir çalışmada, Struthiomimus'a atfedilmiş bulunan elli ayak kemiği, stres kırığı belirtileri açısından incelenmiştir, ancak hiçbir veri bulunamamıştır.[33]
Struthiomimus, başından beri yatay bir duruşa sahip olduğu düşünülen ilk teropodlardan biriydi. 1916'da Osborn, hayvanın kasıtlı olarak yükseltilmiş bir kuyrukla tasvir edilmesine izin verdi.[2] Bu yeni görünüm, dinozorun adının atıfta bulunduğu devekuşu gibi modern uçamayan kuşları çok fazla andıran bir resim yarattı, ancak bu resim çok daha sonra tüm teropodlar için kabul edilecekti.
Beslenme

Struthiomimus'un beslenme alışkanlıkları hakkında çok fazla tartışma yapılmıştır. Düz kenarlı gagası nedeniyle, Struthiomimus bir omnivor olduğu düşünülebilir. Bazı teoriler, onun bir kıyı sakini olabileceğini ve süzerek beslenen bir canlı olabileceğini öne sürmektedir.[18] Bazı paleontologlar, etçil olma olasılığının daha yüksek olduğunu, çünkü farklı bir etçil teropod grubu içinde sınıflandırıldığını belirtti.[3][34] Bu teori hiçbir zaman göz ardı edilmedi, ancak dinozoru tanımlayan ve adlandıran Osborn, muhtemelen ağaçlardan, çalılardan ve diğer bitkilerden tomurcukları ve sürgünleri yediğini,[16] ön ayaklarını dalları kavramak için kullandığını ve uzun boynunun da açık bir şekilde özel öğeleri seçebilecek durumda olduğunu belirtmiştir. Bu otçul diyet, ellerinin sıra dışı yapısı ile daha da desteklenir. İkinci ve üçüncü parmaklar eşit uzunluktadır, bağımsız olarak işlev göremeyen bu parmaklar muhtemelen tek bir birim olarak deri ile birbirine bağlıydı. Omuz kuşağının yapısı, kolun yüksek bir şekilde kaldırılmasına izin vermemektedir ve düşük bir erişim için ayarlanmıştır.
Struthiomimus'un eli, kavrama ve eğilme konusunda tamamen esneyememektedir. Bu durum, dinozorun elinin, dalları veya eğrelti otu yapraklarını omuz hizasına getirmek için bir "kanca" veya "kelepçe" olarak kullanıldığını gösterir.[19] Ancak, bu uyarlamalar bunun yerine kanat tüyü desteği için kullanılmış olabilir.[20]
Hız
Struthiomimus'un bacakları (arka uzuvları) uzun, güçlü ve bir devekuşu gibi hızlı koşmaya çok uygun görünmektedir. Struthiomimus'un varsayılan hızı, aslında, Gorgosaurus, Daspletosaurus, Dromaeosaurus gibi yırtıcılara karşı ana savunma hareketidir (Sıkıştırıldıklarında arka ayaklarındaki pençelerle saldırabilme olasılıkları da olmasına rağmen). Struthiomimus'un 50 ve 80 kilometre/saat (31 ve 50 mph) arasındaki hızlarda koşabildiği tahmin ediliyor.[35]
Dağılım ve yaş
S. altus'un fosil kalıntıları, Geç Kretase döneminin Kampaniyen evresinde 78 ila 77 milyon yıl öncesine tarihlenen Oldman Formasyonu'ndan kesin olarak bilinmektedir.[9] S. altus'tan daha uzun, daha ince ellere sahip olarak ayrışan daha genç bir tür (henüz adlandırılmamış), 69 ila 67.5 milyon yıl önce oluşmuş olan Horseshoe Kanyon Formasyonu ve aşağı Lance Formasyonu'nda bulunan birkaç örnekten bilinmektedir. (Erken Maastrihtiyen ).[9]
Konuyla ilgili yayınlar
- Russell (1969). "A new specimen of Stenonychosaurus from the Oldman Formation (Cretaceous) of Alberta". Canadian Journal of Earth Sciences. 6 (4): 595-612. doi:10.1139/e69-059.
- Cranfield, I. (2004). Dinozorlar ve Diğer Tarih Öncesi Yaratıkların Resimli Dizini (s. 30–33). Greenwich Sürümleri.0-86288-662-7ISBN'si 0-86288-662-7 .
- Reisdorf (2012). "Re-evaluating Moodie's Opisthotonic-Posture Hypothesis in fossil vertebrates. Part I: Reptiles - The taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen Archipelago (Jurassic, Germany)". Palaeobiodiversity and Palaeoenvironments. 92: 119-168. doi:10.1007/s12549-011-0068-y.
Dış bağlantılar
 Wikimedia Commons'ta Struthiomimus ile ilgili çoklu ortam belgeleri bulunur
Wikimedia Commons'ta Struthiomimus ile ilgili çoklu ortam belgeleri bulunur
Kaynakça
- ^ Barrett (2005). "The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria)". Palaeontology. 48 (2): 347-358. doi:10.1111/j.1475-4983.2005.00448.x.
- ^ a b c d e Osborn (1917). "Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus" (PDF). Bulletin of the American Museum of Natural History. 35: 733-771. 20 Temmuz 2007 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 31 Ağustos 2021.Osborn, Henry Fairfield (1917). "Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus" 20 Temmuz 2007 tarihinde Wayback Machine sitesinde arşivlendi. (PDF). Bulletin of the American Museum of Natural History. 35: 733–771. Kaynak hatası: Geçersiz
<ref>etiketi: "HFO17" adı farklı içerikte birden fazla tanımlanmış (Bkz: ) - ^ a b Russell D (1972). "Ostrich dinosaurs from the Late Cretaceous of Western Canada". Canadian Journal of Earth Sciences. 9 (4): 375-402. doi:10.1139/e72-031.Russell D (1972). "Ostrich dinosaurs from the Late Cretaceous of Western Canada". Canadian Journal of Earth Sciences. 9 (4): 375–402. Bibcode:1972CaJES...9..375R 30 Temmuz 2020 tarihinde Wayback Machine sitesinde arşivlendi.. doi:10.1139/e72-031.
- ^ McFeeters (2016). "A new ornithomimid theropod from the Dinosaur Park Formation of Alberta, Canada". Journal of Vertebrate Paleontology. 36 (6): e1221415. doi:10.1080/02724634.2016.1221415.
- ^ Parks (1926). "Struthiomimus brevetertius - A new species of dinosaur from the Edmonton Formation of Alberta". Transactions of the Royal Society of Canada. Series 3. 20 (4): 65-70.
- ^ Parks (1928). "Struthiomimus samueli, a new species of Ornithomimidae from the Belly River Formation of Alberta". University of Toronto Studies, Geology Series. 26: 1-24.
- ^ Parks (1933). "New species of dinosaurs and turtles from the Upper Cretaceous formations of Alberta". University of Toronto Studies, Geological Series. 34: 1-33.
- ^ Glut, D., 1997, Dinosaurs - The Encyclopedia. McFarland Press, Jefferson, NC. 1076 pp
- ^ a b c d e Claessens (2015). "A redescription of Ornithomimus velox Marsh, 1890 (Dinosauria, Theropoda)". Journal of Vertebrate Paleontology. 36: e1034593. doi:10.1080/02724634.2015.1034593.
- ^ a b c Longrich (2008). "A new, large ornithomimid from the Cretaceous Dinosaur Park Formation of Alberta, Canada: Implications for the study of dissociated dinosaur remains". Palaeontology. 51 (4): 983-997. doi:10.1111/j.1475-4983.2008.00791.x.Longrich, N (2008). "A new, large ornithomimid from the Cretaceous Dinosaur Park Formation of Alberta, Canada: Implications for the study of dissociated dinosaur remains". Palaeontology. 51 (4): 983–997. doi:10.1111/j.1475-4983.2008.00791.x.
- ^ Marsh (1892). "Notice of new reptiles from the Laramie Formation". American Journal of Science. Series 3. 43 (257): 449-453. doi:10.2475/ajs.s3-43.257.449. 31 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 31 Ağustos 2021.
- ^ Farlow, J.O., 2001, "Acrocanthosaurus and the maker of Comanchean large-theropod footprints", In: Tanke, Carpenter, Skrepnick and Currie (eds). Mesozoic Vertebrate Life: New Research Inspired by the Paleontology of Philip J. Currie. pp. 408-427
- ^ Aaron (2016). "[2015] A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada". Cretaceous Research. 58: 108-117. doi:10.1016/j.cretres.2015.10.004.
- ^ "Ornithomimus altus". Predatory Dinosaurs of the World. New York: Simon & Schuster. 1988. ss. 387-389. ISBN 978-0-671-61946-6.
- ^ Récords y curiosidades de los dinosaurios Terópodos y otros dinosauromorfos. İspanya: Larousse. 2016. s. 269.
- ^ a b "Genus Ornithomimus". Predatory Dinosaurs of the World. New York: Simon & Schuster. 1988. ss. 384-394. ISBN 978-0-671-61946-6.
- ^ a b "Theropods, Including Birds". Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press. 2005. ss. 367-397. ISBN 978-0-253-34595-0.Currie, Philip J. (2005). "Theropods, Including Birds". In Currie, Phillip J.; Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press. pp. 367–397. ISBN 978-0-253-34595-0.
- ^ a b "Ornithomimosauria". The Dinosauria. 2nd. Berkeley: University of California Press. 2004. ss. 137-150. ISBN 978-0-520-24209-8.
- ^ a b Nicholls (1985). "Structure and function of the pectoral girdle and forelimb of Struthiomimus altus (Theropoda: Ornithomimidae)". Palaeontology. 28: 643-677.
- ^ a b Zelenitsky (2012). "Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins". Science. 338 (6106): 510-514. doi:10.1126/science.1225376. PMID 23112330.Zelenitsky, D. K.; Therrien, F.; Erickson, G. M.; Debuhr, C. L.; Kobayashi, Y.; Eberth, D. A.; Hadfield, F. (2012). "Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins". Science. 338 (6106): 510–514. Bibcode:2012Sci...338..510Z 1 Eylül 2021 tarihinde Wayback Machine sitesinde arşivlendi.. doi:10.1126/science.1225376. PMID 23112330.
- ^ Marsh (1890). "Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs". American Journal of Science. 39 (233): 418-426. doi:10.2475/ajs.s3-39.233.418. 26 Şubat 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 31 Ağustos 2021.
- ^ Marsh (1895). "On the affinities and classification of the dinosaurian reptiles". American Journal of Science. 50 (300): 483-498. doi:10.2475/ajs.s3-50.300.483. 31 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 31 Ağustos 2021.
- ^ O. C. Marsh. 1896. The dinosaurs of North America. United States Geological Survey, 16th Annual Report, 1894-95 55:133-244
- ^ Baur (1891). "Remarks on the reptiles generally called Dinosauria". The American Naturalist. 25 (293): 434-454. doi:10.1086/275329.
- ^ Russell (1993). "The affinities of a new Theropod from the Alxa Desert, Inner Mongolia, People's Republic of China". Canadian Journal of Earth Sciences. 30 (10–11): 2107-2127. doi:10.1139/e93-183.
- ^ J. A. Gauthier and K. Padian. 1985. Phylogenetic, functional, and aerodynamic analyses of the origin of birds and their flight. In M. K. Hecht, J. H. Ostrom, G. Viohl, and P. Wellnhofer (eds.), The Beginnings of Birds: Proceedings of the International Conference Archaeopteryx, Eichstätt 1984. Freunde des Jura-Museums Eichstätt, Eichstätt 185-197
- ^ F. E. Novas. 1992. The evolution of carnivorous dinosaurs. In J. L. Sanz and A. D. Buscalioni (eds.), The Dinosaurs and Their Environment Biotic: Proceedings of the Second Year of Paleontology in Cuenca. Institute "Juan Valdez", Cuenca, Argentina 126-163
- ^ Sereno (1994). "Early Cretaceous dinosaurs from the Sahara". Science. 266 (5183): 267-271. doi:10.1126/science.266.5183.267. PMID 17771449.
- ^ "Ornithomimosauria". The Dinosauria. 2nd. Berkeley: University of California Press. 2004. ss. 137-150. doi:10.1525/california/9780520242098.003.0008. ISBN 9780520242098.
- ^ R. Barsbold. 1976. K evolyutsii i sistematike pozdnemezozoyskikh khishchnykh dinozavrov [The evolution and systematics of late Mesozoic carnivorous dinosaurs]. In N. N. Kramarenko, B. Luvsandansan, Y. I. Voronin, R. Barsbold, A. K. Rozhdestvensky, B. A. Trofimov & V. Y. Reshetov (eds.), Paleontology and Biostratigraphy of Mongolia. The Joint Soviet-Mongolian Paleontological Expedition, 3:68-75 Transactions
- ^ Sereno (1998). "A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 210 (1): 41-83. doi:10.1127/njgpa/210/1998/41.
- ^ Li Xu, Yoshitsugu Kobayashi, Junchang Lü, Yuong-Nam Lee, Yongqing Liu, Kohei Tanaka, Xingliao Zhang, Songhai Jia and Jiming Zhang (2011). "A new ornithomimid dinosaur with North American affinities from the Late Cretaceous Qiupa Formation in Henan Province of China". Cretaceous Research. 32 (2): 213-222. doi:10.1016/j.cretres.2010.12.004.[]
- ^ Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
- ^ Osmólska H, Roniewicz E (1972). "A new dinosaur, Gallimimus bullatus n. gen.,n. sp. (Ornithomimidae) from the Upper Cretaceous of Mongolia". Paleontol. Polonica. 27: 103-143.
- ^ Paul, regarding his comparative speed estimates, notes that "... just how swift is swift? In hard, precise measure, this can be a real can of worms; for just how fast living animals run is not well known." (Paul, G.S. 1988. Predatory Dinosaurs of the World. New York: Simon & Schuster.)



















