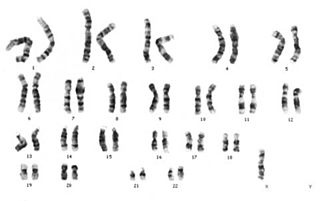
Sitogenetik

Sitogenetik genetiğin bir dalıdır. Sitogenetik aynı zamanda sitolojinin bir parçasıdır. Kromozomların hücre davranışıyla; özellikle mitoz bölünme ve mayoz bölünme sırasındaki davranışlarıyla nasıl bir ilişkili içinde olduğunu inceler.[1] Kullanılan teknikler arasında karyotipleme, G-bantlı kromozomların analizi bulunmaktadır. Floresan in situ hibridizasyon (FISH) ve komparatif genomik hibridizasyon (KGH) gibi teknikler moleküler sitogenetik başlığı altında yer almaktadır.
Tarihi
Başlangıç
Kromozomlar ilk olarak 1842 yılında bitki hücrelerinde çalışmalar yapan Karl Wilhelm von Nägeli tarafından gözlemlenmiştir. Kromozom davranışları semender hücreleri üzerinde çalışmalar yapan Walther Flemming tarafından tanımlanmıştır. Mitoz bölünme 1882 yılında bilim insanı Walther Flemming tarafından keşfedilmiştir . "Sitogenetik" ismi ise 1888 yılında Alman anatomist von Waldeyer tarafından koyulmuştur.
Bir sonraki aşamaya 20. yüzyılın başlarında geçilmiştir Bu geçiş kromozom setinin (karyotip) genlerin taşıyıcısı olarak değerlendirildiği zaman gerçekleşmiştir. Levitsky, karyotipi genetik içeriklerinin aksine vücut kromozomların fenotipik görünümü olarak tanımlayan ilk kişi olduğu varsayılmaktadır.[2][3] "Normal bir diploid insan hücresi kaç kromozom içerir?" sorusuna cevap bulmak için insan karyotipi üzerine yapılan çalışmalar yıllarca devam etmiştir.[4] 1912 yılında Hans von Winiwarter, sperm ana hücresinde (spermatagonyum) 47 kromozom olduğunu; yumurta ana hücresinde (oogonyum) ise 48 kromozom olduğunu bildirdi. Bu bildirmeler XX / XO biyolojik cinsiyet belirleme sistemini ortaya çıkardı.[5] 1922 yılında Theophilus Painter, kromozom sayısının 46 veya 48 olduğu hakkında kesin yargılarda bulunamasa da kromozom sayısını 46 almayı tercih etmiştir.[6] Painter, sonrasında fikrini 46'dan 48'e revize etti. Bunun yanında insanların XX / XY cinsiyet belirleme sistemine sahip olduğu konusunda ısrarlarına devam etmiştir.[7] Kullanılan tekniklerine bakıldığında sonuçlar oldukça dikkat çekici bir etki yaratmıştı. Otuz yıldan fazla bir süre boyunca bilim kitaplarında insan kromozom sayısı 48 kalmıştır. Yeni tekniklerin gelişmesi ve bu hatanın düzelmesi gerekmiştir. Bu görev için Albert Levan'ın laboratuvarında çalışan Joe Hin Tjio[8][9] seçilmiştir. Yapılan araştırmaların doğrultusunda insanın karyotipinin sadece 46 kromozom içerdiği kanıtlanmıştır. Bu araştırmaların bir sonucuna ulaşması ise 1956 yılına kadar devam etmiştir.[10][11][12] Büyük maymunlarda 48 kromozom bulunmaktadır. İnsan kromozomlarından biri olan kromozomu 2; ata kromozomlarının birleşmesi sonucu oluşmuştur. Bu birleşmeden dolayı kromozom sayısı azalmıştır.[13]
Uygulama alanları
McClintock'un mısır bitkisi üzerindeki çalışması
Barbara McClintock kariyerine mısır sitogenetikçisi olarak başlamıştır. McClintock ve Harriet Creighton, 1931 yılında yaptıkları araştırmalarla belirgin kromozomların sitolojik eşleşmesinin (rekombinasyon) genetik özelliklerin rekombinasyonuna bağlı olduğunu göstermiştir. McClintock, Carnegie Enstitüsüde çalışmalarına devam etmiştir. Aynı zaman aralığında mısır bitkisinde kromozom kırılması ve füzyon alevlenmesi mekanizmaları üzerine yaptığı çalışmalarına devam etmiştir. McClintock mısır kromozomu 9'da her zaman aynı gen bölgesinde meydana gelen kromozom kırılma olayını tanımlamıştır. Bu durumu "Ds" veya "ayrışma" lokusu olarak adlandırmıştır.[14]
Drosophila'daki doğal popülasyonlar
Theodosius Dobzhansky ve çalışma arkadaşları, 1930'lu yıllarda bir çalışma yapmışlardır. Bu çalışmada Kaliforniya ve komşu eyaletlerdeki vahşi popülasyonlardan Drosophila pseudoobscura ve D. persimilis'i toplamışlardır. Painter'ın tekniğini[15] kullanarak politen kromozomlarını incelemişlerdir. İncelemeler sonucunda vahşi popülasyonların kromozomal inversiyonlar için çok çeşitli olduğunu keşfetmişlerdir. Bütün sineklerin, taşıdıkları inversiyonlar ne olursa olsun birbirlerine benzer göründükleri gözlemlenmiştir. Bu, şifreli bir polimorfizm örneğidir.
Bu durumun doğal seçilim yüzünden olduğuna ilişkin kanıtlar hızla birikmiştir. İlk defa Dobzhansky, L'Héritier ve Teissier tarafından denenmiş bir yöntem kullanılmıştır. Bu yöntem popülasyonları kafeslerde besleyerek beslenme, üreme ve örneklemeyi mümkün kılmıştır. Canlılarının kaçışını ise önlemiştir. 1951 yalında ise Dobzhansky kitabının üçüncü baskısını yayınlamıştır.[16] Polimorfizmde gözlemlenen; heterozigotların seçici avantajıyla canlı topluluğunda kromozom yapısının değişmediğini kanıtlamıştır.[17][18]
Zambak ve fare
Zambak, mayozun sitolojik incelemesi için tercih edilen bir organizmadır. Bunun sebebi zambak kromozomlarının büyük olmasıdır. Mayozun her morfolojik aşaması mikroskop altında rahatça tanımlanabilir. Hotta ve arkadaşları[19] mayoz bölünmenin profaz aşamasında kemirgenlerin erkek mayotik hücrelerinde çapraz geçişin meydana geldiği varsayılmıştır. Bu varsayım DNA kesilmesi ve onarım sentezinin ortak bir örüntü oluşması için ortam hazırlamıştır. Zambak ve fare gibi filogenetik olarak birbirinden uzak organizmalar arasında ortak bir modelin var olması yazarları bir sonuca ulaştırmıştır. Bu sonuç gelişmiş ökaryotlarda mayotik geçiş organizasyonunun dağıtım açısından evrensel olduğu sonucuna varılmasına yol açmıştır.
Teknikler
Karyotipleme
Rutin kromozom analizi (Karyotipleme), tripsin ve Giemsa boyası, Leishman boyası veya ikisinin bir karışımı kullanılarak metafaz kromozomlarının analizine verilen addır. Yapılan işlemler kromozomlarda eşsiz bant desenleri yaratmaktadır. Bu maddelerin replikasyon zamanlaması ve kromatin paketleme ile ilişkisi vardır. Fakat bu ilişki modellerinin moleküler mekanizması ve nedeni bilinmemektedir.
Sitogenetik laboratuvarlarında birçok kromozom bantlama tekniği kullanılmaktadır. Kinakrin bantlama (K-bantlama), belirli bant desenleri oluşturmak için uygulanan ilk boyama yöntemidir. Bu yöntem bir floresan mikroskobu gerektirmektedir. Fakat Giemsa bantlama (G-bantlama) kadar yaygın olarak kullanılmamaktadır. Ters bantlama (T-bantlama), ısıtma aşaması gerektiren bir bantlama tekniğidir. G-bantlarında ve Q-bantlarında görülen normal siyah beyaz deseni tersine çevirmek üzere kullanılır. T-bantlama spesifik olarak kromozomların uzak uçlarını boyamak için daha yararlıdır. Boyama teknikleri arasında C-bantlama ve çekirdekçikte organize bölge lekeleri (ÇOB lekeleri) bulunmaktadır. C-bantlama ve ÇOB lekeleri kromozomun belirli kısımlarının boyanmasında kullanılır. C-bandı, sentromere yakın olan yapıcı heterokromatini boyar. NOR boyama ise akrosantrik kromozomların uydularını ve saplarını vurgular.
Karyotipleme için kullanılan tekniklerden bir tanesi de yüksek çözünürlüklü bantlama tekniğidir. Bu teknik, kromozomların maksimum yoğunlaşmaya ulaşmadan önce profaz veya erken metafaz aşamasında boyanması işlemini içinde barındırır. Profaz ve erken metafaz aşamalarında, kromozomlar metafaz evresindeki kromozomlardan daha geniştir. Bundan dolayı tüm kromozomlar için gözlemlenebilen bantların sayısı yaklaşık 300'den 450'ye ve 800'e çıkar. Yüksek Çözünürlüklü bantlama geleneksel bantlama teknikleri ile gözlemlenmeyen nadir anormalliklerin tespitinde kullanılır.
Lamların hazırlanması
Kemik iliği, kan, amniyotik sıvı, kordon kanı, tümör ve dokulardan (deri, göbek bağı, karaciğer ve diğer birçok organ dahil) hücreler alınarak standart hücre kültüründe kültürlenirler. Kültürlenme işleminin amacı ise hücrelerin sayısını çoğaltmaktır. Daha sonra hücre kültürüne bir antimitotik (kolşisin, kolsemid) eklenir. Antimitotik, mitozda bölünmeyi durdurarak analiz için mitotik hücre verimini artırır. Hücreler santrifüjlenir. Daha sonra antimitotik, ortamdan uzaklaştırılır. Hücreler hipotonik bir ortama konur. Hipotonik ortam beyaz kan hücrelerinin veya fibroblastların şişmesine neden olur. Slaytlara eklendiğinde kromozomlar yayılırlar. Bu durum kırmızı kan hücrelerini parçalar. Hücrelerin hipotonik çözelti içinde oturmasına izin verilir. Bu işlemden sonra çözeltiye Carnoy tespit sıvısı (3:1 metanolden buzlu asetik asite) eklenir. Yapılan işlem hücreleri öldürür ve beyaz kan hücrelerinin çekirdeklerini sertleştirir. Bu işlem herhangi bir birikintiyi veya kırmızı kan hücresini atmak için tekrar edilir. Hücre süspansiyonu daha sonra numune lamlarına damlatılır. Lamlar bir fırında eskitilir ve beklemeye bırakılır. Birkaç gün bekledikten sonra bantlama ve analiz için hazır hale gelir.
Analiz
Bantlı kromozomların analizi, sitogenetikte uzmanlaşmış klinik laboratuvar uzmanı(KLU(SG)) tarafından mikroskopta gerçekleştirilir. Oluşan mozaikliği kabul edilebilir kılmak için 20 hücrenin analiz edilmesi yeterli bulunur. Sonuçlar özet haline getirilir. Sonuçlar kurul onaylı bir sitogenetik uzmanı tarafından gözden geçirilir. Hastanın geçmişi ve diğer klinik bulgular dikkate alınarak bir yorum yazılır.
Floresan in situ hibridizasyon
Floresan in situ hibridizasyon (FISH), sitogenetik hücre preparatlarına melezlenmesi için floresan etiketli probların kullanılmasına verilen addır.
FISH genel kullanımının yanında aşağıda verilen alanlarda da kullanılabilir:
- kemik iliği yaymaları
- kan yaymaları
- parafine gömülü doku preparatları
- enzimatik olarak ayrışmış doku örnekleri
- kültürlenmemiş kemik iliği
- kültürlenmemiş amniyositler
Lamların hazırlanması
Standart sitogenetik preparatların hazırlanmasına atıfta bulunularak hazırlanmıştır.
Slayt, 2X TSS (tuz, sodyum sitrat) içeren tuz çözeltisi kullanılarak yaşlandırılır. Slaytlar daha sonra etanollü çözeltiye aktarılır. Böylelikle dehidrasyon işlemi uygulanmış olur. Prob karışım eklenir. Örnek DNA ve prob DNA ısıtılmış plaka kullanılarak denatüre edilir. En az 4 saat süreyle yeniden tavlanmaya bırakılır. Slaytlar fazla bağlanmamış sondayı çıkarmak için yıkanır. Sonrasında 4 ', 6-Diamidino-2-fenilindol (DAPI) veya propidyum iyodür ile zıt boyanma işlemleri gerçekleştirilir.
Analiz
FISH numunelerinin analizi, KLU(SG) tarafından floresan mikroskobu ile yapılır. Onkoloji için 200-1.000 arası hücre sayılır ve puanlanır. Bu puanlamanın amacı düşük seviyeli hastalıkları dışlamaktır. Doğuştan gelen sorunlar için 20 metafaz hücresi puanlanır.
Sitogenetiğin geleceği
Yaşanan gelişmeler, standart FISH preparatlarının sonuçlarını saymayı amaçlar. Bundan dolayı otomatik sistemler ve karşılaştırmalı genomik hibridizasyon dizileri, CGH ve Tek nükleotid polimorfizm dizileri gibi sanal karyotipleme teknikleri dahil olmak üzere moleküler sitogenetiklere odaklanmaktadır.
Kaynakça
- ^ Rieger, Rigomar (1976). Glossary of genetics and cytogenetics : classical and molecular. 4th completely rev. ed. Arnd Michaelis, Melvin M. Green. Berlin: Springer-Verlag. ISBN 0-387-07668-9. OCLC 2202589.
- ^ Rodionov, A. V. "Grigorii Andreevich Levitsky (1878–1942)". Russian Journal of Genetics (İngilizce). 45 (11): 1261-1266. doi:10.1134/S1022795409110015. ISSN 1022-7954.
- ^ Haskell, Gordon. "Seedling morphology in applied genetics and plant breeding". The Botanical Review. 27 (3): 382-421. doi:10.1007/bf02860811. ISSN 0006-8101.
- ^ Kottler, M. J. (1974). "From 48 to 46: cytological technique, preconception, and the counting of human chromosomes". Bulletin of the History of Medicine. 48 (4): 465-502. ISSN 0007-5140. PMID 4618149. 22 Mayıs 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mayıs 2021.
- ^ von Winiwarter H (1912). "Études sur la spermatogenese humaine" [Human spermatogenesis studies]. Arch. Biologie (in French). 27 (93): 147–149.
- ^ "Abstracts". The Anatomical Record (İngilizce). 23 (1): 89-132. doi:10.1002/ar.1090230111. ISSN 0003-276X.
- ^ Painter, Theophilus S. "Studies in mammalian spermatogenesis. II. The spermatogenesis of man". Journal of Experimental Zoology (İngilizce). 37 (3): 291-336. doi:10.1002/jez.1400370303. ISSN 0022-104X.
- ^ Wright, Pearce (11 December 2001). "Joe Hin Tjio The man who cracked the chromosome count". The Guardian. Archived from the original on 25 August 2017.
- ^ Saxon, Wolfgang (7 December 2001). "Joe Hin Tjio, 82; Research Biologist Counted Chromosomes". The New York Times. Archived from the original on 12 May 2013.
- ^ Tjio, J. H. (15 Mart 1978). "The chromosome number of man". American Journal of Obstetrics and Gynecology. 130 (6): 723-724. doi:10.1016/0002-9378(78)90337-x. ISSN 0002-9378. PMID 345813. 22 Mayıs 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mayıs 2021.
- ^ Hsu, T. C. (1979). Human and Mammalian Cytogenetics : an Historical Perspective. New York, NY: Springer New York. ISBN 978-1-4612-6159-9. OCLC 852788516.
- ^ "Archived copy 17 Şubat 2011 tarihinde Wayback Machine sitesinde arşivlendi.". Archived from the original on 2011-02-17. Retrieved 2011-03-15. Encyclopædia Britannica, The Human Chromosome
- ^ "Archived copy 9 Ağustos 2011 tarihinde Wayback Machine sitesinde arşivlendi.". from the original on 2011-08-20. Retrieved 2010-05-29. Evolution Pages, Chromosome fusion
- ^ Ravindran, Sandeep (11 Aralık 2012). "Barbara McClintock and the discovery of jumping genes". Proceedings of the National Academy of Sciences of the United States of America. 109 (50): 20198-20199. doi:10.1073/pnas.1219372109. ISSN 1091-6490. PMC 3528533 $2. PMID 23236127. 23 Mayıs 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mayıs 2021.
- ^ Painter, T. S. (22 Aralık 1933). "A NEW METHOD FOR THE STUDY OF CHROMOSOME REARRANGEMENTS AND THE PLOTTING OF CHROMOSOME MAPS". Science (New York, N.Y.). 78 (2034): 585-586. doi:10.1126/science.78.2034.585. ISSN 0036-8075. PMID 17801695. 23 Mayıs 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mayıs 2021.
- ^ Dobzhansky T. 1951. Genetics and the origin of species. 3rd ed, Columbia University Press, New York.
- ^ Dobzhansky T. 1970. Genetics of the evolutionary process. Columbia University Press N.Y.
- ^ [Dobzhansky T.] 1981. Dobzhansky's genetics of natural populations. eds Lewontin RC, Moore JA, Provine WB and Wallace B. Columbia University Press N.Y.
- ^ Hotta, Y.; Chandley, A. C.; Stern, H. (15 Eylül 1977). "Meiotic crossing-over in lily and mouse". Nature. 269 (5625): 240-242. doi:10.1038/269240a0. ISSN 0028-0836. PMID 593319. 23 Mayıs 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Mayıs 2021.