Santral dogma (moleküler biyoloji)
Moleküler biyolojinin santral (merkezi) dogması, biyolojik bir sistem içindeki genetik bilgi akışının bir açıklamasıdır. Orijinal anlamı bu olmasa da, genellikle "DNA RNA'yı, RNA proteini yapar" şeklinde ifade edilir [1] İlk olarak 1957'de Francis Crick[2][3] tarafından ifade edilmiş, 1958'de ise yayınlanmıştır.[4][5]
Francis Crick 1970'te yayınlanan bir Nature makalesinde bunu şöyle ifade etmiştir : "Moleküler biyolojinin santral dogması sıralı bilgilerin ayrıntılı olarak bir molekül zincirinden diğerine transferi ile ilgilenir. Bu tür bilgilerin proteinden ne proteine ne de nükleik aside geri aktarılamayacağını belirtir."[6]

Merkezi dogmanın ikinci bir versiyonu popülerdir ancak yanlıştır. Bu, James Watson tarafından The Molecular Biology of the Gene'nin (1965) ilk baskısında yayınlanan basit DNA → RNA → protein yoludur. Watson'ın versiyonu Crick'inkinden farklıdır çünkü Watson iki aşamalı (DNA → RNA ve RNA → protein) bir süreci merkezi dogma olarak tanımlar.[7] Başlangıçta Crick tarafından belirtildiği gibi dogma bugün geçerliliğini korurken,[6] Watson'ın versiyonu geçerli değildir.[2]
Dogma, en yaygın veya genel durumda, canlı organizmalarda bilgi taşıyan biyopolimerler arasındaki dizi bilgisi transferini anlamak için bir çerçeve sunar. Bu tür biyopolimerlerin 3 ana sınıfı vardır: DNA ve RNA (her ikisi de nükleik asitler) ve protein. Bu üç biyomolekül sınıfı arasında, akla yatkın 3 × 3 = 9 adet doğrudan bilgi aktarımı çeşidi vardır. Dogma bunları 3 grupta sınıflandırır: 3 genel aktarım (çoğu hücrede normal olarak gerçekleştiğine inanılır), 3 özel aktarım (olduğu bilinir, ancak bazı virüsler durumunda veya laboratuvarda yalnızca belirli koşullar altında gerçekleşir) ve 3 bilinmeyen transferler (asla gerçekleşmeyeceği düşünülmektedir). Genel transferler biyolojik bilginin normal akışını tanımlar: DNA, DNA'ya kopyalanabilir (DNA replikasyonu), DNA bilgisi mRNA'ya kopyalanabilir (transkripsiyon) ve mRNA'daki bilgiler bir şablon olarak (translasyon) kullanılarak proteinler sentezlenebilir. Özel transferler şunları açıklar: RNA'dan kopyalanan RNA (RNA replikasyonu), bir RNA şablonu kullanılarak DNA sentezlenir (ters transkripsiyon ) ve mRNA kullanılmadan doğrudan bir DNA şablonundan sentezlenen proteinler. Bilinmeyen transferler şunları tanımlar: bir proteinden kopyalanan bir protein, bir proteinin birincil yapısını şablon olarak kullanarak RNA sentezi ve bir proteinin birincil yapısını şablon olarak kullanarak DNA sentezi - bunların doğal olarak meydana geldiği düşünülmez.[6]
Biyolojik bilgi dizisi
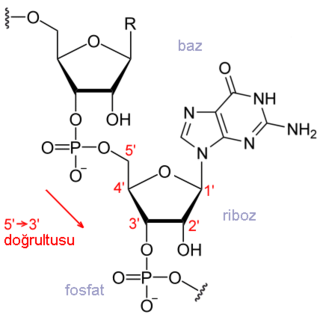
DNA, RNA ve (poli) peptitleri içeren biyopolimerler lineer polimerlerdir (yani: her monomer en fazla iki diğer monomere bağlıdır). Monomerlerinin dizisi bilgiyi etkili bir şekilde kodlar. İdeal olarak, merkezi dogma tarafından açıklanan bilgi transferleri, bir biyopolimerin dizisinin, tamamen orijinal biyopolimerin dizisine bağlı bir diziye sahip başka bir biyopolimerin inşası için bir şablon olarak kullanıldığı, sadık, deterministik transferlerdir. DNA, RNA'ya kopyalandığında, tamamlayıcısı onunla eşlenir. A, G, T ve C DNA kodları sırasıyla U, C, A ve G RNA kodlarına aktarılır. Proteinlerin kodlanması, tabloya göre kodonlar olarak bilinen üçlü gruplar halinde yapılır.

Biyolojik sıralı bilgilerin genel transferleri

DNA replikasyonları
Somatik veya üreme olsun, herhangi bir hücrenin soyuna genetik materyal sağlanacaksa, DNA replikasyonunun gerçekleşmesi gerekir. Replizom adı verilen karmaşık bir protein grubu, bilginin ana iplikten tamamlayıcı yavru iplikçiklere kopyalanmasını gerçekleştirir.[8]
Replisom şunları içerir:
- bir replikasyon çatalı oluşturmak için süper sarmalın yanı sıra çift sarmallı DNA sarmalını çözen helikaz [8]
- Helikaz tarafından ayrlan DNA zincirinin yeniden birleşmesini önlemek için çift sarmallı DNA'yı bağlayan SSB proteini [8]
- Replikasyon için bir başlangıç noktası olarak her bir şablon zincirine tamamlayıcı bir RNA primeri ekleyen RNA primazı [8]
- Mevcut şablon zincirini 3' ucundan 5' ucuna kadar okuyan ve yavru zincirin 5' ucundan 3' ucuna yeni tamamlayıcı nükleotitler ekleyen DNA polimeraz III [8]
- RNA primerlerini çıkaran ve onları DNA ile değiştiren DNA polimeraz I [8]
- Sürekli bir zincir oluşturmak için iki Okazaki fragmanını fosfodiester bağlarıyla birleştiren DNA ligazı [8]
Bu süreç ökaryotik hücrelerde tipik olarak hücre döngüsünün S fazı sırasında gerçekleşir.
Transkripsiyon

Transkripsiyon, DNA'nın bir bölümünde yer alan bilgilerin bir araya getirilmiş bir haberci RNA (mRNA) parçası biçiminde kopyalandığı süreçtir. İşlemi kolaylaştıran enzimler arasında RNA polimeraz ve transkripsiyon faktörleri yer alır . Ökaryotik hücrelerde transkripsiyon işleminde oluşturulan ilk zincir pre-mRNA'dır (öncül mRNA). Bu mRNA'nın translasyonunun gerçekleşebilmesi için öncül mRNA'nın işlenmesi gerekmektedir. öncül mRNA'yı translasyona hazırlayan işlemler bütünü post-transkripsiyonel modifikasyonlar olarak isimlendirilir. Bunlar; ön mRNA zincirine bir 5' başlık ve bir poli-A kuyruğunun eklenmesini içerir . Bazı durumlarda, öncül mRNA'da intronların çııkarılması sürecinde alternatif ekleme-çıkarma (splicing) denilen bir işlem meydana gelir ve herhangi bir tek mRNA'nın üretebileceği proteinlerin çeşitliliğini arttırır. Tüm transkripsiyon sürecinin (öncül mRNA zincirinin üretimiyle başlayan) ürünü, olgun bir mRNA zinciridir.
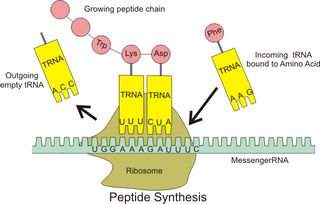
Translasyon
Olgun mRNA, translasyonda görev almak üzere bir ribozoma doğru yol alır. Çekirdeği bulunmayan prokaryotik hücrelerde, transkripsiyon ve translasyon süreçleri net bir ayrılma olmaksızın birbirine bağlıdır. Ökaryotik hücrelerde ise, transkripsiyon bölgesi (hücre çekirdeği) genellikle translasyon bölgesinden (sitoplazma) bariz bir şekilde farklıdır. Bu farklılık nedeniyle transkripsiyonda üretilen mRNA'nın, üretildiği organel olan çekirdekten ribozomlarla bağlanabileceği sitoplazmaya taşınması gerekmektedir. Ribozom, genellikle bir AUG (adenin - urasil - guanin) ile başlayan mRNA üçlü kodonlarını veya ribozom bağlanma bölgesinin aşağısındaki başlatıcı metiyonin kodonunu okur . Başlatma faktörleri ve uzama faktörleri kompleksleri, amino asitleri açillenme tepkimeleri ile bağlamış bulunan transfer RNA'ları (tRNA'lar) ribozom-mRNA kompleksine getirir ve mRNA'daki kodonu tRNA üzerindeki anti-kodonla eşleştirir. Her tRNA, sentezlenen polipeptit zincirine eklenecek uygun amino asit kalıntısını taşır. Amino asitler büyüyen peptit zincirine bağlandıkça, zincir doğru konformasyona katlanmaya başlar. Çeviri, bir UAA, UGA veya UAG üçlüsü olabilen bir durdurma kodonu ile sona erer.
mRNA, olgun proteinin doğasını belirtmek için tüm bilgileri içermez. Ribozomdan salınan, yeni sentezlenmiş polipeptit zinciri, genellikle nihai ürün ortaya çıkmadan önce ek işleme maruz kalır. Bunlar da post-translasyonel modifikasyonlar olarak adlandırılırlar. Bu modifikasyonlardan birincisi, doğru katlama işlemidir ki karmaşık olmakla beraber proteinin işlevi açısından hayati önem taşır. Doru katlanma o denli önemlidir ki birçok proteinin ürün formunu kontrol etmek için şaperon proteinlere ihtiyaç duyulur. Bazı proteinler, sonrasında, kendi peptit zincirlerinden bazı iç kısımları keserek, boşluğu sınırlayan serbest uçları birleştirir. Atılan bu bölümlere inteins adı verilir. Diğer proteinler, ekleme yapılmadan birden çok bölüme ayrılmalıdır. Bazı polipeptit zincirlerinin çapraz bağlanması gerekir ve diğerleri işlevsel hale gelmeden önce haem (heme) gibi kofaktörlere bağlanmalıdır.
Biyolojik sıralı bilgilerin özel transferleri
Ters transkripsiyon
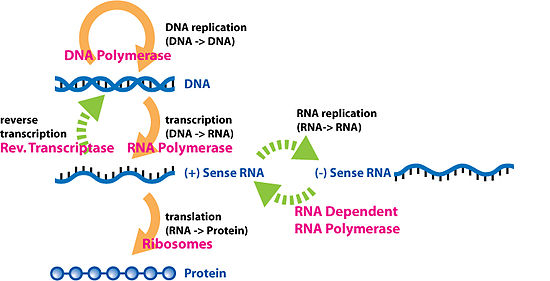
Ters transkripsiyon, RNA'dan DNA'ya bilgi aktarımıdır (normal transkripsiyonun tersi). Bunun HIV gibi retrovirüsler durumunda olduğu kadar ökaryotlarda, retrotranspozonlar ve telomer sentezi durumunda meydana geldiği bilinmektedir. RNA'dan gelen genetik bilginin yeni DNA'ya kopyalandığı süreçtir. Bu sürece dahil olan enzim ailesine Ters Transkriptaz denir.
RNA replikasyonu
RNA replikasyonu, bir RNA'nın diğerine kopyalanmasıdır. Birçok virüs bu şekilde çoğalır. RNA'yı yeni RNA'ya kopyalayan enzimler RNA'ya bağımlı RNA polimerazlar olarak adlandırılırlar. Bu enzimler birçok ökaryotik hücrede de bulunur ve oralarda, çoğunlukla, RNA susturma işleminde görev alırlar.[9]
Bir RNA dizisinin bir protein kompleksi ve bir "kılavuz RNA" tarafından değiştirildiği RNA düzenlemesi, RNA'dan RNA'ya transfer olarak da görülebilir.
DNA'dan proteine doğrudan çeviri
DNA'dan proteine doğrudan translasyon, hücre içermeyen bir sistemde (yani bir test tüpünde), ribozomlar içeren ancak sağlam hücreler içermeyen E. coli özütleri kullanılarak gösterilmiştir. Bu hücre fragmanları, diğer organizmalardan (örneğin, fare veya kurbağa) izole edilen tek sarmallı DNA şablonlarından proteinleri sentezleyebilmektedir ve neomisin'in bu etkiyi arttırdığı bulunmuştur. Bununla birlikte, bu çeviri mekanizmasının özellikle genetik koda karşılık gelip gelmediği açık değildir.[10][11]
Teoride açıkça kapsanmayan bilgi transferleri
Post-translasyonel Modifikasyonlar (Protein sentezi sonrası değişiklikler)
Proteinler, amino asit dizileri nükleik asit zincirleri şablon olarak kullanılarak sentezlendikten sonra uygun enzimler tarafından düzenlenebilirler. Bu, protein-dizisini-etkileyen-protein tarzında bir bilgi olmasına rağmen, santral (merkezi) dogma tarafından açıkça kapsanmamıştır. İki alanın ilişkili kavramlarının birbiriyle çok fazla ilgisi olduğu açık pek fazla örnek yoktur.
İnteinler
Bir intein, bir protein ribozomdan çıktıktan sonra kendisini kesebilen ve ana protein "omurgasının" yaptığı şekilde bir peptit bağı ile geri kalan kısımları yeniden birleştirebilen bir "parazitik" amino asit dizisidir (protein parçasıdır). Bu durum, bir proteinin, DNA'sı tarafından orijinal olarak kodlanan diziye kıyasla kendi birincil dizisini değiştirmesini mümkün kılar. Ek olarak, çoğu intein, intein nükleotid dizisini içermeyen ana genin bir kopyasını bulabilen hedeflenen endonükleaz veya HEG alanı içerir. İntein içermeyen kopya ile temas halinde, HEG alanı, DNA çift sarmallı kopma onarım mekanizmasını başlatır. Bu işlem, intein sekansının orijinal kaynak genden intein içermeyen gene kopyalanmasına neden olur. Bu, DNA dizisini doğrudan düzenleyen ve dizinin kalıtsal yayılımını artıran bir protein örneğidir.
Metilasyon
DNA'nın metilasyon durumlarındaki varyasyon, gen ekspresyon seviyelerini önemli ölçüde değiştirebilir. Metilasyon varyasyonu genellikle DNA metilazların etkisiyle meydana gelir. Değişiklik kalıtsal olduğunda, epigenetik olarak kabul edilir. Bilgi durumundaki değişiklik kalıtsal olmadığında, somatik bir epitip olur . Etkili bilgi içeriği, bir proteinin veya proteinlerin DNA üzerindeki etkileri vasıtasıyla değiştirilmiştir, ancak birincil DNA dizisi değiştirilmemiştir.
Prionlar
Prionlar, belirli amino asit dizilerinin özel konformasyonlarındaki proteinleridir. Aynı amino asit dizisine sahip, ancak fonksiyonel olarak önemli veya organizma için zararlı olan farklı bir konformasyona sahip diğer protein moleküllerinde konformasyonel değişiklikler yaparak konakçı hücrelerde kendilerini çoğaltırlar. Protein prion katlanmasına dönüştürüldüğünde, işlevi değişir. Buna karşılık, bilgiyi yeni hücrelere iletebilir ve bu dizinin daha işlevsel moleküllerini alternatif prion formuna yeniden yapılandırabilir. Mantarlardaki bazı prion türlerinde bu değişim sürekli ve doğrudandır; bilgi akışı Proteindir → Protein.
Alain E. Bussard ve Eugene Koonin gibi bazı bilim insanları, prion aracılı kalıtımın moleküler biyolojinin santral dogmasını ihlal ettiğini savundular.[12][13] Bununla birlikte, Rosalind Ridley, Molecular Pathology of the Prion'da (2001) şöyle yazmıştır: "Prion hipotezi, moleküler biyolojinin santral dogmasına -protein üretmek için gerekli bilginin nükleik asidin nükleotid dizisinde kodlandığına- aykırı değildir. proteinlerin çoğaldığını iddia etmez. Aksine, protein moleküllerinin içinde biyolojik işlevlerine katkıda bulunan bir bilgi kaynağı olduğunu ve bu bilginin diğer moleküllere aktarılabileceğini iddia eder." [14]
Doğal genetik mühendisliği
James A. Shapiro, bu örneklerin bir üst kümesinin doğal genetik mühendisliği olarak sınıflandırılması gerektiğini ve santral dogmayı tahrif etmek için yeterli olduğunu savunuyor. Shapiro, görüşü için saygılı bir duruşma almış olsa da, eleştirmenleri, merkezi dogmayı okumasının Crick'in amaçladığı ile uyumlu olduğuna ikna olmuş değil.[15][16]
Dogma teriminin kullanımı
Crick, What Mad Pursuit adlı otobiyografisinde , dogma kelimesini seçmesi ve bunun neden olduğu bazı sorunlar hakkında şunları yazmıştı:
"Bu fikre santral dogma adını vermemin sanırım iki nedeni var. Dizi hipotezinde zaten açık olan hipotez kelimesini kullanmıştım ve ayrıca bu yeni varsayımın daha merkezi ve daha güçlü olduğunu belirtmek istedim. . . . Anlaşıldığı üzere, dogma kelimesinin kullanılması neredeyse değerinden daha fazla soruna neden oldu. Yıllar sonra Jacques Monod bana, şüpheye yer bırakmayacak bir inanç olan dogma kelimesinin doğru kullanımını anlamamış göründüğüme dikkat çekti. Bunu belirsiz bir şekilde anladım ama tüm dini inançların temelsiz olduğunu düşündüğümden, bu kelimeyi dünyanın çoğu gibi değil, kendim hakkında düşündüğüm şekilde kullandım yani bir hipotezin makul olmasına rağmen, çok az doğrudan deneysel desteğe sahip olması anlamında."
Benzer şekilde Horace Freeland Judson, The Eighth Day of Creation'da [Yaradılışın Sekizinci Günü] şunları ifade etmektedir[17]
"Aklım, dogmanın makul bir kanıtı olmayan bir fikir olduğundaydı. Anlıyor musunuz? !" Ve Crick sevinçle kükredi. "Sadece dogmanın ne anlama geldiğini bilmiyordum . Ve ben buna 'Santral Hipotez' de diyebilirdim, ya da - bilirsiniz. Demek istediğim buydu. Dogma sadece bir slogandı."
Weismann bariyeri ile karşılaştırma

1892'de August Weismann tarafından önerilen Weismann bariyeri, gamet üreten "ölümsüz" eşey hücre soyları (germ plazması) ile "tek kullanımlık" somatik hücreler arasında ayrım yapar. Kalıtsal bilgi yalnızca eşey hücresinden somatik hücrelere geçer (yani somatik mutasyonlar kalıtsal değildir). Bu, DNA'nın rolünün veya yapısının keşfinden önce, merkezi dogmayı öngörmez, ancak moleküler olmayan terimlerle de olsa gen merkezli yaşam görüşünü tahmin eder.[18][19]
Konuyla ilgili yayınlar
- Bussard AE (August 2005). "A scientific revolution? The prion anomaly may challenge the central dogma of molecular biology". EMBO Reports. 6 (8). ss. 691-4. doi:10.1038/sj.embor.7400497. PMC 1369155 $2. PMID 16065057.
- Baker, Harry F. (2001). Molecular Pathology of the Prions (Methods in Molecular Medicine). Humana Press. 0-89603-924-2
- Li JJ, Biggin MD (March 2015). "Gene expression. Statistics requantitates the central dogma". Science. 347 (6226). ss. 1066-7. Bibcode:2015Sci...347.1066L. doi:10.1126/science.aaa8332
 . PMID 25745146.
. PMID 25745146. - Piras V, Tomita M, Selvarajoo K (2012). "Is central dogma a global property of cellular information flow?". Frontiers in Physiology. Cilt 3. s. 439. doi:10.3389/fphys.2012.00439. PMC 3505008 $2. PMID 23189060.
- Robinson VL (2009). "Rethinking the central dogma: noncoding RNAs are biologically relevant". Urologic Oncology. 27 (3). ss. 304-6. doi:10.1016/j.urolonc.2008.11.004. PMID 19414118.
Dış bağlantılar
- Merkezi 10 Kasım 2014 tarihinde Wayback Machine sitesinde arşivlendi. Dogmanın Detaylandırılması – Scitable: By Nature eğitimi
- RIKEN'den Central Dogma Animasyonu 29 Temmuz 2021 tarihinde Wayback Machine sitesinde arşivlendi. - NatureDocumentaries.org
- "Moleküler Biyolojinin Merkezi Dogması"na meydan okumalar üzerine tartışma 25 Ekim 2014 tarihinde Wayback Machine sitesinde arşivlendi.
- Müzikal bir benzetme kullanarak merkezi dogmanın açıklaması 20 Aralık 2014 tarihinde Wayback Machine sitesinde arşivlendi.
- A. Andrei tarafından "Francis Harry Compton Crick (1916–2004)" Embriyo Projesi Ansiklopedisi'nde 19 Ekim 2014 tarihinde Wayback Machine sitesinde arşivlendi.
Kaynakça
- ^ "Deciphering the Genetic Code: Marshall Nirenberg". Office of NIH History. June 2010. 17 Mart 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 2 Mart 2012.
- ^ a b "60 years ago, Francis Crick changed the logic of biology". PLOS Biology. 15 (9): e2003243. September 2017. doi:10.1371/journal.pbio.2003243. PMC 5602739 $2. PMID 28922352.
- ^ "CSHL Archives Repository | On Protein Synthesis". libgallery.cshl.edu (İngilizce). 24 Eylül 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 13 Kasım 2018.
- ^ "On Protein Synthesis". Symposia of the Society for Experimental Biology, Number XII: The Biological Replication of Macromolecules. Cambridge University Press. 1958. ss. 138-163.
- ^ Crick (1958). "On protein synthesis". Symposia of the Society for Experimental Biology. Symposia on the society for Experimental biology number XII: The Biological Replication of Macromolecules. 12. PMID 13580867. 8 Mart 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Temmuz 2021.
- ^ a b c "Central dogma of molecular biology" (PDF). Nature. 227 (5258): 561-3. August 1970. doi:10.1038/227561a0. PMID 4913914. 26 Ocak 2020 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 29 Temmuz 2021.
- ^ "Sandwalk: Basic Concepts: The Central Dogma of Molecular Biology". sandwalk.blogspot.com. 15 Ocak 2007. 20 Ocak 2007 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Mart 2018.
- ^ a b c d e f g "SnapShot: The replisome". Cell. 141 (6): 1088-1088.e1. June 2010. doi:10.1016/j.cell.2010.05.042. PMC 4007198 $2. PMID 20550941.
- ^ "RNA-dependent RNA polymerases, viruses, and RNA silencing". Science. 296 (5571): 1270-3. May 2002. doi:10.1126/science.1069132. PMID 12016304.
- ^ "Denatured DNA as a direct template for in vitro protein synthesis". Proceedings of the National Academy of Sciences of the United States of America. 54 (3): 880-6. September 1965. doi:10.1073/pnas.54.3.880. PMC 219759 $2. PMID 4955657.
- ^ ."Polypeptide synthesis directed by DNA as a messenger in cell-free polypeptide synthesis by extreme thermophiles, Thermus thermophilus HB27 and Sulfolobus tokodaii strain 7". Journal of Biochemistry. 131 (6): 849-53. June 2002. doi:10.1093/oxfordjournals.jbchem.a003174. PMID 12038981.
- ^ "A scientific revolution? The prion anomaly may challenge the central dogma of molecular biology". EMBO Reports. 6 (8): 691-4. August 2005. doi:10.1038/sj.embor.7400497. PMC 1369155 $2. PMID 16065057.
- ^ "Does the central dogma still stand?". Biology Direct. 7: 27. August 2012. doi:10.1186/1745-6150-7-27. PMC 3472225 $2. PMID 22913395.
- ^ Baker, Harry F., (Ed.) (2001). "What Would Thomas Henry Huxley Have Made of Prion Diseases?". Molecular Pathology of the Prions. Methods in Molecular Medicine. Humana Press. ss. 1-16. ISBN 0-89603-924-2.
- ^ Wilkins (January 2012). "(Review) Evolution: A View from the 21st Century". Genome Biology and Evolution. 4 (4): 423-426. doi:10.1093/gbe/evs008. PMC 3342868 $2.
- ^ Moran (May–June 2011). "(Review) Evolution: A View from the 21st Century". Reports of the National Center for Science Education. 32.3 (9): 1-4. 15 Eylül 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Ekim 2012.
- ^ "Chapter 6: My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?!". The Eighth Day of Creation: Makers of the Revolution in Biology. 25th anniversary. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. 1996. ISBN 978-0-87969-477-7.
- ^ "From DNA- to NA-centrism and the conditions for gene-centrism revisited". Biology & Philosophy. 29 (1): 55-69. January 2014. doi:10.1007/s10539-013-9393-z.
- ^ Biology's Second Law: Homeostasis, Purpose, and Desire. Beyond Mechanism: Putting Life Back Into Biology. Rowman and Littlefield. 2013. s. 192. ISBN 978-0-7391-7436-4. 29 Temmuz 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Temmuz 2021.
Where Weismann would say that it is impossible for changes acquired during an organism's lifetime to feed back onto transmissible traits in the germ line, the CDMB now added that it was impossible for information encoded in proteins to feed back and affect genetic information in any form whatsoever, which was essentially a molecular recasting of the Weismann barrier.
| Alanlar, disiplinler |
|  |
|---|---|---|
| Enstitüler |
| |
| Teoriler, kavramlar | ||
| Tarihî | ||
| İlgili konular |
| |