Sanger dizilemesi
Sanger dizilemesi, in vitro DNA replikasyonu sırasında DNA polimeraz tarafından zincir sonlandırıcı dideoksinükleotidlerin seçici bir şekilde dahil edilmesine dayanan bir DNA dizileme yöntemidir.[1][2] İlk olarak 1977'de Frederick Sanger ve meslektaşları tarafından geliştirildikten sonra, yaklaşık 40 yıldır en yaygın kullanılan dizileme yöntemi haline geldi. İlk olarak 1986 yılında Applied Biosystems tarafından ticarileştirildi.[3] Daha yakın zamanlarda, daha yüksek hacimli Sanger dizilemesi, özellikle büyük ölçekli, otomatik genom analizleri için "Gelecek-Nesil" dizileme yöntemleriyle değiştirildi. Bununla birlikte, Sanger yöntemi, daha küçük ölçekli projeler ve Gelecek-Nesil sonuçların doğrulanması için yaygın olarak kullanılmaktadır. Yine de, kısa okumalı dizileme teknolojilerine (Illumina gibi) göre, >500 nükleotidlik DNA dizisi okumaları üretebilme avantajına sahiptir.
Yöntem

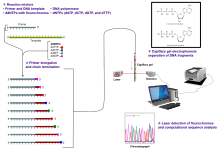
Klasik zincir sonlandırma yöntemi, tek sarmallı bir DNA şablonu, bir DNA primeri, bir DNA polimeraz, normal deoksinükleotidtrifosfatlar (dNTPs) ve modifiye di-deoksinükleotidtrifosfatlar (ddNTPs) gerektirir, bunlardan ikincisi DNA sarmalının uzamasını sonlandırır. Bu zincir sonlandırıcı nükleotidler, iki nükleotid arasında bir fosfodiester bağının oluşumu için gerekli olan bir 3'-OH grubundan yoksundur, bu da DNA polimerazın, modifiye bir ddNTP dahil edildiğinde DNA'nın uzamasını durdurmasına neden olur. DdNTP'ler, otomatik dizileme makinelerinde tespit için radyoaktif olarak veya floresan olarak etiketlenebilir.
DNA örneği, standart deoksinükleotidlerin dördünü (dATP, dGTP, dCTP ve dTTP) ve DNA polimerazı içeren dört ayrı dizileme reaksiyonuna bölünmüştür. Her reaksiyona dört dideoksinükleotidden (ddATP, ddGTP, ddCTP veya ddTTP) sadece biri eklenirken, eklenen diğer nükleotidler sıradan olanlardır. Tam diziyi hala kopyalarken yeterli fragmanın üretilmesine izin vermek için deoksinükleotid konsantrasyonu (örn. 0,5 mM dTTP: 0,005 mM ddTTP), karşılık gelen dideoksinükleotidinkinden yaklaşık 100 kat daha yüksek olmalıdır (ancak ddNTP konsantrasyonu aynı zamanda istenen dizi uzunluğuna da bağlıdır).[2] Daha mantıklı bir sıraya koyarsak, bu süreçte dört ddNTP'nin tümünü test etmek için dört ayrı reaksiyona ihtiyaç vardır. Bağlı primerden şablon DNA uzatma turlarının ardından, elde edilen DNA fragmanları ısıyla denatüre edilir ve jel elektroforezi kullanılarak boyuta göre ayrılır. 1977 tarihli orijinal yayında,[2] baz çiftli ssDNA döngülerinin oluşumu, bazı yerlerde bantları çözmede ciddi zorlukların bir sebebiydi. Bu sıklıkla denatüre edici bir poliakrilamid-üre jeli kullanılarak gerçekleştirilir ve dört reaksiyonun her biri dört ayrı şeritten birinde (A, T, G, C şeritleri) yürütülür. DNA bantları daha sonra otoradyografi veya UV ışığı ile görselleştirilebilir ve DNA dizisi doğrudan X-ışını filmi veya jel görüntüsünden okunabilir.

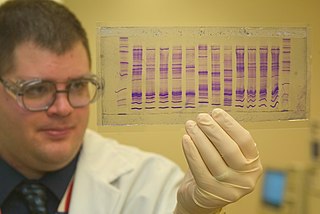
Sağdaki görüntüde, X-ışını filmi jele maruz bırakıldı ve koyu bantlar, farklı uzunluklardaki DNA fragmanlarına karşılık geliyor. Bir şeritteki koyu bir bant, bir dideoksinükleotidin (ddATP, ddGTP, ddCTP veya ddTTP) dahil edilmesinden sonra zincir sonlandırmasının sonucu olan bir DNA parçasını belirtir. Aşağıdan yukarıya dört şerit arasındaki farklı bantların göreceli konumları daha sonra DNA dizisini okumak için kullanılır.

Zincir sonlandırma dizilemesinin teknik varyasyonları arasında radyoaktif etiketleme için radyoaktif fosfor içeren nükleotidlerle etiketleme veya 5'ucunda bir floresan boya ile etiketlenmiş bir primer kullanılması yer alır. Boya astar dizilemesi, daha hızlı ve daha ekonomik analiz ve otomasyon için optik bir sistemde okumayı kolaylaştırır. Leroy Hood ve floresan etiketli ddNTP'ler ve primerlerin iş arkadaşları[4][5] tarafından yapılan sonraki geliştirme, otomatikleştirilmiş, yüksek verimli DNA dizilimi için zemin hazırladı.

Zincir sonlandırma yöntemleri, DNA dizilimini büyük ölçüde basitleştirmiştir. Örneğin, zincir sonlandırmaya dayalı kitler, dizileme için gerekli reaktifleri içeren, önceden bölünmüş ve kullanıma hazır olan ticari olarak mevcuttur. Sınırlamalar arasında primerin DNA'ya spesifik olmayan bağlanması, DNA dizisinin doğru okunmasını ve dizinin doğruluğunu etkileyen DNA sekonder yapıları bulunur.
Boya-sonlandırıcı dizilemesi

Boya sonlandırıcı dizilemesi, etiketli primer yöntemindeki gibi dört reaksiyon yerine tek bir reaksiyonda dizilemeye izin veren zincir sonlandırıcı ddNTP'lerin etiketlemesini kullanır. Boya sonlandırıcı dizilemede, dört dideoksinükleotid zincir sonlandırıcıdan her biri, her biri farklı dalga boylarında ışık yayan floresan boyalarla etiketlenir.
Daha uygunluğu ve hızı sayesinde, boya sonlandırıcı dizileme artık otomatik dizilemede temel dayanaktır. Sınırlamaları, boya etiketli zincir sonlandırıcıların DNA fragmanına dahil edilmesindeki farklılıklardan kaynaklanan boya etkilerini içerir, bu da kılcal elektroforezden sonra elektronik DNA dizisi izleme kromatogramında eşit olmayan tepe yükseklikleri ve şekilleriyle sonuçlanır.
Bu sorun, "boya lekelerini" ortadan kaldırmak için yöntemlerin yanı sıra, dahil etme değişkenliğini en aza indiren modifiye edilmiş DNA polimeraz enzim sistemleri ve boyalarının kullanılmasıyla ele alınmıştır. Boya sonlandırıcı dizileme yöntemi, otomatik yüksek verimli DNA dizisi analizörleriyle birlikte, Gelecek-Nesil Dizileme'nin tanıtımına kadar dizileme projelerinin büyük çoğunluğunda kullanıldı.
Otomasyon ve örnek hazırlama

Otomatik DNA dizileme cihazları (DNA sıralayıcılar), tek bir grupta 384'e kadar DNA örneğini sıralayabilir. Toplu çalıştırmalar günde 24 defaya kadar gerçekleşebilir. DNA sıralayıcıları kılcal elektroforez kullanarak şeritleri boyuta (veya uzunluğa) göre ayırır, boya floresansını tespit eder ve kaydeder ve flüoresan tepe izleme kromatogramları olarak çıktı verileri. Örnekleri sıralayıcıya yüklemeden önce, bir tampon solüsyonunda örneklerin dizileme reaksiyonları (ısıl döngü ve etiketleme), temizlenmesi ve yeniden süspansiyonu ayrı ayrı gerçekleştirilir. Bir dizi ticari ve ticari olmayan yazılım paketi, düşük kaliteli DNA izlerini otomatik olarak kesebilir. Bu programlar her zirvenin kalitesini puanlar ve düşük kaliteli temel zirveleri kaldırır (genellikle dizinin sonlarında bulunur). Bu tür algoritmaların doğruluğu, bir insan operatör tarafından yapılan görsel incelemeden daha düşüktür, ancak büyük sıralı veri setlerinin otomatik olarak işlenmesi için yeterlidir.
Zorluklar
Sanger yöntemiyle DNA dizilemesinin yaygın zorlukları, primer bağlanması nedeniyle dizinin ilk 15-40 bazında düşük kaliteyi ve 700-900 bazdan sonra dizileme izlerinin kalitesinin bozulmasını içerir. Phred gibi temel arama yazılımı, tipik olarak, dizilerin düşük kaliteli bölgelerinin kırpılmasına yardımcı olmak için bir kalite tahmini sağlar.[6][7]
DNA fragmanlarının dizilemeden önce klonlandığı durumlarda, ortaya çıkan dizi, klonlama vektörünün parçalarını içerebilir. Bunun aksine, PCR tabanlı klonlama ve piroz sıraya dayalı yeni nesil dizileme teknolojileri genellikle klonlama vektörlerini kullanmaktan kaçınır. Son zamanlarda, Ampliseq ve SeqSharp gibi tek adımlı Sanger dizileme (kombine amplifikasyon ve dizileme) yöntemleri, klonlama veya önceden amplifikasyon olmadan hedef genlerin hızlı dizilemesine izin veren geliştirilmiştir.[8][9]
Mevcut yöntemler, tek bir reaksiyonda yalnızca nispeten kısa (300-1000 nükleotid uzunluğunda) DNA parçalarını doğrudan sıralayabilir. Bu boyut sınırının üzerindeki DNA fragmanlarının sıralanmasının önündeki ana engel, uzunlukları sadece bir nükleotid kadar farklılık gösteren büyük DNA fragmanlarını çözmek için yetersiz ayırma gücüdür.
Mikroakışkan Sanger dizilemesi
Sanger dizileme adımlarının (termal döngü, örnek saflaştırma ve kapiler elektroforez) nanolitre ölçekli örnek hacimleri kullanılarak wafer-ölçekli bir çip üzerine entegre edildiği, DNA dizilemesi için bir çip üzerinde laboratuvar uygulamasıdır. Bu teknoloji, Sanger dizileme adımlarını entegre ederek ve otomatikleştirerek geleneksel Sanger yönteminin (örn. pahalı reaktiflerin yüksek tüketimi, pahalı ekipmanlara güvenme, personel yoğun manipülasyonlar, vb.) birçok önemli eksikliğini ortadan kaldırırken, uzun ve doğru dizi okumaları üretir.
Modern başlangıcında, yüksek verimli genom dizilimi, genomun küçük tek sarmallı parçalara bölünmesini ve ardından parçaların Polimeraz Zincir Reaksiyonu ile amplifikasyonunu içerir. Sanger yöntemini benimseyen her DNA fragmanı, flüoresan olarak etiketlenmiş bir dideoksi zincirini sonlandıran nükleotidin dahil edilmesiyle geri döndürülemez bir şekilde sonlandırılır, böylece her biri uzunluğu bir baz ile farklılık gösteren ve terminal bazında baza özgü bir floresan etiket taşıyan bir DNA fragmanları üretir. Güçlendirilmiş baz merdivenleri daha sonra, parçaların sıralı bir dizisini sağlayan floresan etiketli ssDNA parçalarının otomatik, yerinde "bitiş çizgisi" tespiti ile Kapiler Dizi Elektroforezi ile ayrılır. Bu dizi okumaları daha sonra bilgisayar, tam olarak birleştirildikten sonra tam genomik diziye benzeyen üst üste binen veya bitişik dizilemeler ("contigs" olarak adlandırılır) halinde birleştirilir.[10]
Sanger yöntemleri, yaklaşık 800bp'lik okuma uzunluklarına ulaşır (tipik olarak zenginleştirilmemiş DNA ile 500-600bp). Sanger yöntemlerinde daha uzun okuma uzunlukları, diğer dizileme yöntemlerine göre, özellikle genomun tekrarlayan bölgelerinin dizilenmesi açısından önemli avantajlar sergilemektedir. Kısa okunan dizi verilerinin zorluğu, özellikle yeni genomların (de novo) dizilemesinde ve yüksek düzeyde yeniden düzenlenmiş genom bölümlerinin, tipik olarak kanser genomlarında veya yapısal varyasyon sergileyen kromozom bölgelerinde görülenlerin dizilemesinde bir sorundur.[11]
Mikroakışkan dizileme teknolojilerinin uygulamaları
DNA dizilemesinin diğer yararlı uygulamaları arasında tek nükleotid polimorfizm saptama, tek sarmallı konformasyon polimorfizm heterodubleks analizi ve kısa tandem tekrar analizi yer alır. DNA parçalarının boyut ve/veya konformasyon farklılıklarına göre çözümlenmesi, genomun bu özelliklerinin incelenmesinde en kritik adımdır.[10]
Cihaz tasarımı
Sıralama çipi, 100 mm çapında üç cam gofret ve bir polidimetilsiloksan membrandan oluşan dört katmanlı bir yapıya sahiptir. Reaksiyon odaları ve kapiler elektroforez kanalları, termal olarak bağlanmış en üstteki iki cam wafer (hangi cihaz öğelerinin mikrofabrike olduğu) arasında oyulmuştur. Üç boyutlu kanal ara bağlantıları ve mikrovalfler, PDMS ve alt manifold cam yonga plakası tarafından oluşturulur.
Cihaz, her biri Sanger dizileme adımlarına karşılık gelen üç işlevsel birimden oluşur. Termal-döngü ünitesi, entegre dirençli sıcaklık dedektörü, mikrovalfler ve bir yüzey ısıtıcısı olan 250 nanolitre reaksiyon odasıdır. Reaktifin en üstteki tüm cam katman ile alt cam-PDMS katmanı arasındaki hareketi, 500 µm çaplı geçiş delikleri aracılığıyla gerçekleşir. Termal-döngüden sonra, reaksiyon karışımı yakalama/saflaştırma bölmesinde saflaştırmaya tabi tutulur ve ardından kapiler elektroforez bölmesine enjekte edilir. CE ünitesi, 65 μm genişliğindeki dönüşlerle kompakt bir geri dönüş düzenine katlanan 30 cm'lik bir kapilerden oluşur.
Dizileme kimyası
Termal-döngü
Termal-döngü reaksiyon odasında, boya sonlandırıcı sekanslama reaktifi, şablon DNA ve primerler, termal-döngü odasına yüklenir ve 35 döngü boyunca termal döngü yapılır (95 °C'de 12 saniye ve 60 °C'de 55 saniye).
Arıtma
Yüklenen reaksiyon karışımı (uzatma parçaları, şablon DNA ve fazla dizileme reaktifi içerir), yakalama çıkışı ve giriş delikleri arasına uygulanan 33 Volt/cm'lik bir elektrik alanı aracılığıyla 30 °C'de bir yakalama/saflaştırma bölmesinden geçirilir. Örneğin sürüldüğü yakalama jeli, bir poliakrilamid matrisine kovalent olarak bağlanmış 40 μM oligonükleotidden (primerleri tamamlayıcı) oluşur. Uzatma fragmanları, jel matris ile hareketsizleştirilir ve fazla primer, şablon, serbest nükleotitler ve tuzlar, yakalama atık portu yoluyla elüte edilir. Yakalama jeli, uzatma parçalarını serbest bırakmak için 67-75 °C'ye ısıtılır.
Kapiler elektroforez
Uzatma fragmanları, 125-167-V/cm alan aracılığıyla elektroforeze tabi tutuldukları kapiler-elektrofez bölmesine enjekte edilir.
Platformlar
Apollo 100 platformu,[12] ilk iki Sanger sıralama adımını (termal-döngü ve arıtma) tam otomatik bir sistemde birleştirir. Üretici, örneklerin ve reaktiflerin sisteme yüklenmesinin ardından üç saat içinde kapiler elektroforez için hazır olduğunu iddia ediyor. Apollo 100 platformu, mikrolitre altı reaktif hacmi gerektirir.
Diğer dizileme teknikleriyle karşılaştırmalar
| Teknoloji | Şerit sayısı | Enjeksiyon hacmi (nL) | Analiz süresi | Ortalama okuma uzunluğu | Çıktı (analiz dahil;Mb/sa ) | Jel dökülmesi | Şerit takibi |
|---|---|---|---|---|---|---|---|
| Döşeme jeli | 96 | 500-1000 | 6-8 saat | 700 bp | 0,0672 | Evet | Evet |
| Kapiler dizi elektroforezi | 96 | 1-5 | 1-3 saat | 700 bp | 0.166 | Hayır | Hayır |
| Mikroçip | 96 | 0.1-0.5 | 6-30 dakika | 430 bp | 0.660 | Hayır | Hayır |
| 454/Roche FLX (2008) | <0.001 | 4 saat | 200-300 bp | 20-30 | |||
| Illumina/Solexa (2008) | 2-3 gün | 30-100 bp | 20 | ||||
| ABI/SOLiD (2008) | 8 gün | 35 bp | 5-15 | ||||
| Illumina MiSeq (2019) | 1-3 gün | 2x75-2x300 bp | 170-250 | ||||
| Illumina NovaSeq (2019) | 1-2 gün | 2x50-2x150 bp | 22.000-67.000 | ||||
| Ion Torrent Ion 530 (2019) | 2,5-4 saat | 200-600 bp | 110-920 | ||||
| BGI MGISEQ-T7 (2019) | 1 gün | 2x150 bp | 250.000 | ||||
| Pacific Biosciences SMRT (2019) | 10-20 saat | 10–30 kb | 1.300 | ||||
| Oxford Nanopore MinIon (2019) | 3 gün | 13–20 kb[15] | 700 |
Yüksek verimli dizilimin nihai amacı, düşük maliyetli ve uzun okuma uzunlukları elde etmede son derece verimli sistemler geliştirmektir. Her bir elektroforetik ayırmanın daha uzun okuma uzunlukları, de novo DNA dizilemesi ile ilişkili maliyeti ve belirli bir artıklıkta DNA bileşenlerini dizilemek için gereken şablon sayısını önemli ölçüde azaltır. Mikroakışkanlar daha hızlı, daha ucuz ve daha kolay dizi montajına izin verebilir.[10]
Kaynakça
- ^ Sanger F; Coulson AR (Mayıs 1975). "A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase". J. Mol. Biol. 94 (3): 441-8. doi:10.1016/0022-2836(75)90213-2. PMID 1100841.
- ^ a b c Sanger F; Nicklen S; Coulson AR (Aralık 1977). "DNA sequencing with chain-terminating inhibitors". Proc. Natl. Acad. Sci. U.S.A. 74 (12): 5463-7. Bibcode:1977PNAS...74.5463S. doi:10.1073/pnas.74.12.5463. PMC 431765 $2. PMID 271968.
- ^ Adams, Jill U. (2008). "DNA Sequencing Technologies". Nature Education. 19 Ekim 2008 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Ekim 2019.
- ^ Smith LM; Sanders JZ; Kaiser RJ; Hughes, Peter; Dodd, Chris; Connell, Charles R.; Heiner, Cheryl; Kent, Stephen B. H.; Hood, Leroy E. (1986). "Fluorescence detection in automated DNA sequence analysis". Nature. 321 (6071): 674-9. Bibcode:1986Natur.321..674S. doi:10.1038/321674a0. PMID 3713851.
- ^ Smith LM; Fung S; Hunkapiller MW; Hunkapiller TJ; Hood LE (Nisan 1985). "The synthesis of oligonucleotides containing an aliphatic amino group at the 5' terminus: synthesis of fluorescent DNA primers for use in DNA sequence analysis". Nucleic Acids Res. 13 (7): 2399-412. doi:10.1093/nar/13.7.2399. PMC 341163 $2. PMID 4000959.
- ^ "Phred - Quality Base Calling". 15 Aralık 2001 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Şubat 2011.
- ^ Ledergerber, C; Dessimoz, C (2011). "Base-calling for next-generation sequencing platforms". Briefings in Bioinformatics. 12 (5): 489-97. doi:10.1093/bib/bbq077. PMC 3178052 $2. PMID 21245079.
- ^ Murphy, K.; Berg, K.; Eshleman, J. (2005). "Sequencing of genomic DNA by combined amplification and cycle sequencing reaction". Clinical Chemistry. 51 (1): 35-39. doi:10.1373/clinchem.2004.039164. PMID 15514094.
- ^ Sengupta, D. .; Cookson, B. . (2010). "SeqSharp: A general approach for improving cycle-sequencing that facilitates a robust one-step combined amplification and sequencing method". The Journal of Molecular Diagnostics. 12 (3): 272-277. doi:10.2353/jmoldx.2010.090134. PMC 2860461 $2. PMID 20203000.
- ^ a b c Kan, Cheuk-Wai; Fredlake, Christopher P.; Doherty, Erin A. S.; Barron, Annelise E. (1 Kasım 2004). "DNA sequencing and genotyping in miniaturized electrophoresis systems". Electrophoresis. 25 (21–22): 3564-3588. doi:10.1002/elps.200406161. PMID 15565709.
- ^ a b Morozova, Olena; Marra, Marco A (2008). "Applications of next-generation sequencing technologies in functional genomics". Genomics. 92 (5): 255-64. doi:10.1016/j.ygeno.2008.07.001. PMID 18703132.
- ^ Microchip Biologies Inc. Apollo 100 28 Ağustos 2008 tarihinde Wayback Machine sitesinde arşivlendi.
- ^ Sinville, Rondedrick; Soper, Steven A (2007). "High resolution DNA separations using microchip electrophoresis". Journal of Separation Science. 30 (11): 1714-28. doi:10.1002/jssc.200700150. PMID 17623451.
- ^ Kumar, Kishore; Cowley, Mark; Davis, Ryan (2019). "Next-Generation Sequencing and Emerging Technologies". Seminars in Thrombosis and Hemostasis. 45 (7): 661-673. doi:10.1055/s-0039-1688446. ISSN 0094-6176. PMID 31096307.
- ^ Tyson (2018). "MinION-based long-read sequencing and assembly extends the Caenorhabditis elegans reference genome". Genome Research. 28 (2): 266-274. doi:10.1101/gr.221184.117. ISSN 1088-9051. PMC 5793790 $2. PMID 29273626.