
DNA replikasyonu veya DNA ikileşmesi, tüm organizmalarda meydana gelen ve DNA kopyalayarak kalıtımın temelini oluşturan biyolojik bir süreçtir. Süreç, bir adet çift iplikli DNA molekülüyle başlar ve iki özdeş DNA'nın oluşumuyla son bulur. Orijinal çift iplikli DNA'nın her ipliği, tamamlayıcı ipliğin üretiminde kalıp görevi görür. Hücresel proofreading ve hata kontrol mekanizmaları replikasyonun neredeyse hatasız gerçekleşmesini sağlar.
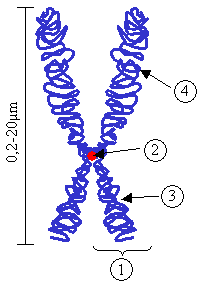
Kromozom, ; DNA'nın "histon" proteinleri etrafına sarılmasıyla, yoğunlaşarak oluşturduğu, canlılarda kalıtımı sağlayan genetik birimlerdir. Kromozomlar mikrometre boyutunda olup hücre bölünmesinin metafaz aşamasında ışık mikroskobu ile görüntülenebilmektedirler.
Mutasyon ya da değişinim, bir canlının genomu içindeki DNA ya da RNA diziliminde meydana gelen kalıcı değişmelerdir. Mutasyona sahip bir organizma ise mutant olarak adlandırılır.

Nükleik asitler, bütün canlı hücrelerde ve virüslerde bulunan, nükleotid birimlerden oluşmuş polimerlerdir. En yaygın nükleik asitler deoksiribonükleik asit (DNA) ve ribonükleik asit (RNA)'dır. İnsan kromozomlarını oluşturan DNA milyonlarca nükleotitten oluşur. Nükleik asitlerin başlıca işlevi genetik bilgi aktarımını sağlamaktır.
Bakteriyel konjugasyon hücre teması yoluyla bakteriler arasında genetik malzeme aktarımıdır. Transformasyon ve transdüksiyon gibi bu da bir yatay gen transferi mekanizmasıdır.

DNA onarımı, DNA moleküllerindeki hataları onarım mekanizmalarını tanımlamaktadır. İnsan hücrelerinde metabolik aktiviteler ve çevresel faktörler sonucu günde 1 milyon hücrenin zarar görmesi olasıdır. Bu etkenler, DNA'nın yapısını ve dahası diğer nesillere aktarılan genetik bilgiyi değiştirebilirler. Bu değişimler yararlı olabileceği gibi, ölümcül sonuçlara neden olabilecek kadar da zararlı olabilir. Bu yüzden, bütün canlı hücreleri, evrim süreçleri boyunca nesillere değişmeden aktarılması gereken DNA molekülünü koruma mekanizmaları geliştirmişlerdir.

Telomer, ökaryotik doğrusal kromozomların uçlarında bulunan, herhangi bir gen kodlamayan, özelleşmiş heterokromatin yapılarıdır.

İkileşme çatalı ya da Replikasyon çatalı replikasyon bölgesinde bulunan "Y" şeklindeki kromozom bölgesidir.

Okazaki parçaları, DNA ikileşmesi sırasında nispeten kısa ve parçalar halinde bulunan zincirlere denir.
Kesintili zincir, döşeme iplik ya da Lagging strand, DNA ikileşmesi sırasında birçok başlangıç noktasına gerek duyan ve Okazaki parçalarının oluştuğu zincirdir.
Kesintisiz zincir, öncü iplik ya da Leading strand DNA zincirinde ikileşme çatalı açıldıkça, kesintili zincirin karşısında yer alan zincirdir.
Prokaryotik hücrelerin DNA ikileşmesinde, ikili sarmal açılır ve sentezin başladığı yer olan ikileşme çatalı oluşur. Proteinler açılan sarmalı kararlı kılar ve ikileşme çatalının önünde oluşan sarılma gerilimini hafifletirler. Sentez, kalıp boyunca belirli bölgelerden RNA Primazın, DNA Polimeraz III'ün polimerizasyonu başlatabileceği serbest 3'-OH ucunu sağlayan kısa bir RNA parçasını sentezlemesiyle başlar. İkili sarmalın antiparalel yapısından dolayı polimeraz III, kesintili zincirde 5'-3' yönünde sürekli DNA sentezi yapar. Çatalın solunda DNA sentezi 5'-3' yönünde kesintisiz olarak devam eder. Kesintili zincir denen karşı zincirde kısa Okazaki parçaları sentezlenir ve bu parçalar daha sonra DNA ligaz ile birleştirilir. DNA Polimeraz I, RNA primerini uzaklaştırır ve yerine DNA sentezler, ortaya çıkan polinükleotidler DNA ligaz ile birleştirilir. Böylece sentezi tamamlanan iki yeni çift dallı DNA molekülü birbirinden ayrılr ve biri atasal hücrede kalırken diğeri oğul hücreye gider.
Ökaryotlarda DNA ikileşmesi, oldukça karmaşık bir işlem olup, DNA sentezindeki bazı faktörlerin nasıl işlediği hala tam olarak çözümlenememiştir.

Crosover veya krossing over ya da parça değişimi mayoz bölünmenin profaz I evresinde görülen, çift halde bulunan kromozomların yaptığı parça değişimine verilen addır. Bunun sonucunda genetik rekombinasyon meydana gelir. Yani farklı kromozomlarda bulunan genlerin alelleri birbiriyle yer değiştirir.
Moleküler biyolojide bir baz çifti, birbirine ters doğrultuda iki DNA veya RNA zinciri üzerinde bulunan, biribirine hidrojen bağları ile bağlanmış iki nükleobazdır. Standart Watson-Crick baz eşleşmesinde, adenin (A), timin (T) ile, guanin de sitozin ile bir baz çifti oluşturur. RNA içinde olan baz çiftlerinde timin'in yerini urasil (U) alır. Watson-Crick tipi olmayan ve alternatif hidrojen bağlarıyla meydana gelmiş baz çiftleri de oluşabilir, özellikle RNA'da; bunlara Hoogsteen baz çiftlerinde de rastlanır.

DNA polimeraz, DNA replikasyonunu sağlayan bir enzimdir. Bu enzimler bir DNA ipliğini kalıp olarak kullanır, onu okuyup, onun boyunca deoksiribonükleotitlerin polimerizasyonunu katalizler. Yeni polimerleşmiş molekül kalıp ipliği tamamlayıcıdır ve kalıp ipliğin eski eşi ile aynı yapıya sahiptir.
Genetik çeşitlenme veya rekombinasyon, genetik materyalin bir zincirinin kırılması ve sonrasında farklı bir DNA molekülüne katılmasıyla oluşan süreçtir. Ökaryotlarda çeşitlilik genellikle mayoz sırasında, kromozom çiftleri arasındaki kromozomal parça değişimiyle meydana gelir. Bu süreç döllerin ebeveynlerinden farklı gen dizilimlerine sahip olmasına ve daha önce olmayan yeni alleler üretmesine öncülük eder.

Biyokimyada bir ters transkriptaz veya RNA'ya bağımlı DNA polimeraz, tek iplikli bir RNA molekülü okuyup tek iplikli DNA üreten bir DNA polimeraz enzimidir. Bu enzim, ayrıca, RNA tek iplikli cDNA şeklinde okunduktan sonra çift iplikli DNA oluşmasında da görev alır. Normal transkripsiyon DNA'dan RNA sentezidir; dolayısıyla ters transkripsiyon bu sürecin tersidir.

Homolog rekombinasyon, benzer veya aynı dizilere sahip DNA iplikleri arasında nükleotit dizilerinin birbiriyle yer değiştirdiği bir genetik rekombinasyon tipidir. Bu süreç sırasında DNA birkaç kere kesilir, sonra da birleştirilir. Homolog rekombinasyon, DNA'daki çift iplikli kırıkların hatasız tamirinde kullanılmanın yanı sıra, mayoz sırasında krosover yoluyla yeni DNA dizi bileşimlerinin (kombinezonlarının) oluşumunu da sağlar. DNA'daki yeni bileşimler genetik varyasyonlar oluşturur. Genetik varyasyonlar yeni, bir olasılıkla yararlı olabilecek alel kombinasyonlarıdır, bunların üreyen canlı topluluklarda oluşmaları, bu değişiklikleri taşıyan bireylerin değişen çevresel şartlara evrimsel adaptasyon göstermelerini sağlar.
Meselson-Stahl deneyi, DNA ikileşmesinin yarı korumalı olduğu hipotezini destekleyen, Matthew Meselson ve Franklin Stahl tarafından yapılmış bir deneydi. Yarı korumalı ikileşmenin (replikasyonun) anlamı, DNA sarmalının iki ipliği ikileşince, meydana gelen her bir çift iplikli DNA sarmalındaki ipliklerden birinin orijinal sarmaldan geldiği, öbürünün ise yeni sentezlenmiş olduğudur. 1958'de yayımlanan bu deneyin sonuçları moleküler biyolojinin en önemli deneyleri arasında yer alır.
