Ornithomimosauria
| Ornitomimozorlar | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 7 Ornithomimosaur cinsinin kolajı, sol üstten saat yönünde: Gallimimus, Anserimimus, Ornithomimus, Deinocheirus, Harpymimus, Struthiomimus ve "Gallimimus mongoliensis" | |||||||||||||||
| Biyolojik sınıflandırma | |||||||||||||||
| |||||||||||||||
| Altgruplar[9] | |||||||||||||||
| Sinonimler | |||||||||||||||
| |||||||||||||||
Ornithomimosauria, ornitomimozorlar ("kuş taklitçisi kertenkeleler") veya devekuşu dinozorları,[10] günümüz devekuşuna yüzeysel bir benzerlik gösteren theropod dinozorlardır. Laurasia'nın Kretase Dönemi'nden (şimdi Asya, Avrupa ve Kuzey Amerika), ayrıca Afrika ve muhtemelen Avustralya'dan hızlı, dz.omnivor veya otçul dinozorlardı.[11] Grup ilk olarak Erken Kretase'de ortaya çıktı ve Geç Kretase'ye kadar varlığını sürdürdü. Grubun ilkel üyeleri arasında kolları 2,4 m (8 fit) uzunluğa ulaşan Nqwebasaurus, Pelecanimimus, Shenzhousaurus, Hexing ve Deinocheirus bulunur. Ornithomimidae ailesinin üyeleri olan daha gelişmiş türler arasında Gallimimus, Struthiomimus ve Ornithomimus bulunur. Paul Sereno gibi bazı paleontologlar, esrarengiz alvarezsauridleri ornithomimosaurların yakın akrabaları olarak kabul eder ve onları Ornithomimoidea üst familyasında bir araya getirir (aşağıdaki sınıflandırmaya bakınız).
Tanım


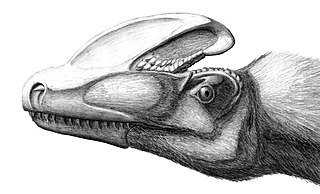
Ornithomimosaurların kafatasları, nispeten uzun ve ince boyunların üzerinde, iri gözlü ve küçüktü. Taksonun en temel üyeleri (Pelecanimimus ve Harpymimus gibi) küçük dişlere sahip bir çeneye sahipken, daha sonraki ve daha türetilmiş türlerin dişsiz bir gagası vardı.[12] Ön uzuvlar ("kollar") uzun ve inceydi ve güçlü pençeleri vardı. Arka uzuvlar uzun ve güçlüydü, uzun bir ayak ve kısa, güçlü ayak parmakları toynak benzeri pençelerle sona eriyordu. Ornithomimosaurlar muhtemelen tüm dinozorların en hızlıları arasındaydı. Diğer coelurosaurlar gibi, ornithomimosaurian postu da pullu olmaktan ziyade tüylüydü.
Tüyler
Ornithomimus edmontonicus'tan tüylerin açık bir kanıtı bilinmektedir ki bunların tüy izlerini koruyan çok sayıda örneği vardır.[13] Deinocheirus ve Pelecanimimus'un da tüylü olduğu, ilkinin bir pygostil varlığı nedeniyle ve daha sonra olası izlenimler nedeniyle (aksi takdirde kollajen lifleri olarak kabul edilir) olduğu tahmin edilmiştir.[14] Ornithomimidlerin Pennaraptora'da görülen pennaceous tüylere sahip olup olmadığı konusunda ise tartışma vardır.[15] Aksi takdirde, bilinen bir örnekte çok devekuşu benzeri bir tüy ve tüy aralığı görülmektedir.[16]
Sınıflandırma
1890'da Othniel Charles Marsh tarafından isimlendirilen Ornithomimidae ailesi, başlangıçta bir "megalozor" (orta ila büyük boyutlu theropod dinozorları içeren bir "çöp sepeti taksonu") olarak sınıflandırılmıştı, ancak daha fazla theropod çeşitliliği ortaya çıktıkça, onların diğer theropodlarla olan gerçek ilişkileri çözülmeye başladı ve Coelurosauria'ya taşındılar atandılar. Rinchen Barsbold, ornithomimidlerin diğer dinozorlara kıyasla ayırt edici özelliğini fark ederek, ornithomimidleri 1976'da kendi alt takımları olan Ornithomimosauria içine yerleştirdi. Ornithomimidae ve Ornithomimosauria'nın içeriği, 1990'larda gruplar için Kladistik tanımlar ortaya çıkmaya başladığından, yazardan yazara değişiyordu.
1990'ların başında, Thomas R. Holtz Jr. gibi önde gelen paleontologlar, arctometatarsal ayaklı theropodlar arasında yakın bir ilişki olduğunu öne sürdüler; yani, üst ayak kemiklerinin birbirine 'kıstırıldığı', koşmaya bir uyarlama olan iki ayaklı dinozorlar. Holtz (1994), Arctometatarsalia soyunu "arctometatarsalia pesini geliştiren ilk theropod ve onun onun soyundan gelenlerin tümü" olarak tanımladı. Bu grup Troodontidae, Tyrannosauroidea ve Ornithomimosauria'yı içeriyordu. Holtz (1996, 2000) daha sonra bu tanımı dal tabanlı "Ornithomimus ve tüm theropodlar, kuşlardan çok Ornithomimus ile daha yakın bir ortak ataya sahip olacak şekilde" rafine etti. Daha sonra, tüm arctometatarsal dinozorlarının doğal bir grup oluşturduğu fikri, Holtz dahil çoğu paleontolog tarafından terk edildi, çünkü çalışmalar, tyrannosauridlerin ve troodontidlerin diğer coelurosaur gruplarıyla ornithomimosaurlardan daha yakından ilişkili olduğunu göstermeye başladı. Arctometatarsalia'nın katı tanımı Ornithomimus'a dayandığından, bu dalın geniş tanımları altında Ornithomimosauria adıyla gereksiz hale geldi ve Arctometatarsalia adı çoğunlukla terk edildi.
Paleontolog Paul Sereno, 2005 yılında, "Ornithomimiformes" dalını önerdi ve onları Passer domesticus'tan çok Ornithomimus edmontonicus'a daha yakın türler olarak tanımladı. Ornithomimosauria'yı çok daha dar bir anlamda yeniden tanımladığı için, tercih ettiği terminolojide Ornithomimosauria ve Alvarezsauridae kardeş gruplarını içeren yeni bir terim gerekli hale getirildi - daha önce ikincisi birincinin içindeydi. Ancak bu konsept sadece Sereno'nun Web sitesinde yer aldı ve henüz resmi olarak geçerli bir isim olarak yayınlanmadı. "Ornithomimiformes", çok benzer bir tanıma sahip olduğu için içerik olarak Holtz'un Arctometatarsalia'sıyla aynıydı. "Ornithomimiformes" daha yeni grup olsa da, Sereno Arctometatarsalia'nın öncelikli olması gerektiği fikrini reddetti, çünkü eski adın anlamı Holtz tarafından çok radikal bir şekilde değiştirildi.[17]
Filogeni

Ornithomimosauria, kuşlardan çok Ornithomimus'a daha yakın olan tüm dinozorların dal tabanlı grubu için ve daha kısıtlayıcı anlamlarda çeşitli şekillerde kullanılmıştır. Kapsayıcı bir tanım kabul edilirse alvarezsauridlerin Ornithomimosauria kapsamına girme olasılığı ortaya çıktığında, daha özel anlam popülerlik kazanmaya başladı. Başka bir dal, Ornithomimiformes, Sereno (2005) tarafından (Ornithomimus velox > Bayağı serçe) olarak tanımlanmıştır ve alvarezsauridlerin veya başka bir grubun ornitomimosaurların maniraptoranlardan daha yakın akrabaları olduğu tespit edildiğinde Ornithomimosauria'nın daha kapsamlı kullanımının yerini alır ve Ornithomimosauria'yı içerecek şekilde yeniden tanımlanır: Ornithomimus'a alvarezsauridlerden daha yakın dinozorlar. Gregory S. Paul, Ornithomimosauria'nın Deinonychosauria ve Oviraptorosauria'dan daha gelişmiş, ilkel, uçamayan bir kuş grubu olabileceğini öne sürdü.[18]
Aşağıdaki kladogram Yuong-Nam Lee, Rinchen Barsbold, Philip J. Currie, Yoshitsugu Kobayashi, Hang-Jae Lee, Pascal Godefroit, François Escuillié & Tsogtbaatar Chinzorig tarafından yapılan bir analizi takip etmektedir. Analiz 2014'te yayınlandı ve birçok ornithomimosaurian taksonunu içeriyor.[9]
| Coelurosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




Aşağıdaki kladogram, Scott Hartman ve meslektaşları tarafından 2019'da türlerin ve belirsiz örneklerin büyük çoğunluğunu içeren ve yeni bir filogenetik düzenlemeyle sonuçlanan filogenetik analizini takip ediyor.[19]
| Ornithomimosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Aşağıda, ornithomimosaur sınıflandırmasıyla ilgili önceki varsayımlarla daha uyumlu bir analizi gösteren, Serrano-Brañas ve diğerleri (2020), tarafından hazırlanan bir kladogram bulunmaktadır.[20]
| Ornithomimosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiyoloji
Ornithomimosaurlar muhtemelen kalorilerinin çoğunu bitkilerden aldılar. İlkel türler de dahil olmak üzere birçok ornithomimosaurun midelerinde otçulların özelliği olan çok sayıda mide taşı bulunmuştur. Henry Fairfield Osborn, ornithomimosaurların uzun, "tembel hayvan benzeri" kollarının, beslenecekleri dalları aşağı çekmek için kullanılmış olabileceğini öne sürdü; bu, tuhaf, kanca benzeri elleri üzerinde daha fazla çalışma ile desteklenen bir fikirdir.[21] Ornithomimidlerin bolluğu - bunlar Kuzey Amerika'daki en yaygın küçük dinozorlardır - otçulların bir ekosistemde genellikle etoburlardan daha fazla olması nedeniyle, bitki yiyiciler oldukları fikriyle tutarlıdır. Bununla birlikte, hem bitkileri hem de küçük hayvanları yiyen omnivorlar olabilirler.
İki ornithomimosaur cinsinin (Garudimimus ve Ornithomimus) skleral halkaları ile modern kuşlar ve sürüngenler arasındaki karşılaştırmalar, bunların gün boyunca kısa aralıklarla aktif, katemeral olabileceğini göstermektedir.[22]
Sosyal davranış

Ornithomimosaurlar, sürü halindeki yaşam tarzları ile oldukça iyi bilinirler. Ornithomimosaur kemik yataklarının ilk bulgularından bazıları, 1993 yılında Charles W. Gilmore tarafından Iren Dabasu Formasyonu'ndan rapor edilmiştir. Kemik yatağı, gençlerden yetişkin kalıntılarına kadar çok sayıda Archaeornithomimus bireyinden oluşuyordu.[23] Tek bir monospesifik kemik yatağından çok sayıda Sinornithomimus örneği, hatırı sayılır miktarda genç bireyden (14 kişiden 11'i gençti) toplanmıştır ve bu da yırtıcılardan daha fazla koruma için toplu bir davranış olduğunu düşündürür. Gençlerin kayda değer bolluğu, onlarda yüksek bir ölüm oranına veya tüm bir grubun büyük bir toplu ölüm olayının meydana geldiğine ve gençlerde daha fazla duyarlılığa sahip olduğunu gösterir. Ek olarak, Sinornithomimus'un ontogenisi yoluyla tibia-femur oranındaki artış, yetişkinlerde gençlere göre daha yüksek imleç kapasitelerine işaret edebilir.[24] Ayrıca, Sinornithomimus kemik yatağının aksine, Bayshi Tsav bölgesinden yüksek konsantrasyonda ornitomimozor örnekleri, farklı ontogenetik aşamalarda en az beş bireyden oluşan tek bir multitaksik kemik yatağında toplanmıştır. Bu kemik yatağının bireylerinin, en az iki farklı taksonun tanımlanması göz önüne alındığında, tek bir türün stratejik bir sosyal davranışını temsil etmesi olası değildir. Bu düşünceye göre, 10'dan fazla farklı ornithomimosaurian sürüsünden oluşan küçük bir sürünün, yiyecek kaynakları, yuvalama alanları veya başka bir şey bulmak için en uygun alanlarda birlikte seyahat etmesi mümkündür.[25][26]
Tanımlanan cinlerin zaman çizelgesi

Kaynakça
- ^ a b Alifanov, V. R.; Saveliev, S. V. (Kasım 2015). "The most ancient Ornithomimosaur (Theropoda, Dinosauria), with cover imprints from the Upper Jurassic of Russia". Paleontological Journal (İngilizce). 49 (6): 636-650. doi:10.1134/S0031030115060039. ISSN 0031-0301.
- ^ Brownstein CD. (2016) Redescription of Arundel formation Ornithomimosaur material and a reinterpretation of Nedcolbertia justinhofmanni as an "Ostrich Dinosaur": Biogeographic implications. PeerJ Preprints 4:e2308v1 https://doi.org/10.7287/peerj.preprints.2308v1
- ^ Choiniere, J. N.; Forster, C. A.; De Klerk, W. J. (2012). "New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous (Hauteriverian?) Kirkwood Formation in South Africa". Journal of African Earth Sciences. 71–72: 1-17. Bibcode:2012JAfES..71....1C. doi:10.1016/j.jafrearsci.2012.05.005.
- ^ "Walgettosuchus woodwardi". Australian Age of Dinosaurs. 13 Kasım 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Kasım 2020.
- ^ Cuesta E, Vidal D, Ortega F, Shibata M, Sanz JL (2021). "Pelecanimimus (Theropoda: Ornithomimosauria) postcranial anatomy and the evolution of the specialized manus in Ornithomimosaurs and sternum in maniraptoriforms". Zoological Journal of the Linnean Society. in press. doi:10.1093/zoolinnean/zlab013.
- ^ Sereno, P. (2017). "Early Cretaceous ornithomimosaurs (Dinosauria: Coelurosauria) from Africa". Ameghiniana. 54 (5): 576-616. doi:10.5710/AMGH.23.10.2017.3155.
- ^ Jin Liyong, Chen Jun; Pascal Godefroit (2012). "A New Basal Ornithomimosaur (Dinosauria: Theropoda) from the Early Cretaceous Yixian Formation, Northeast China". Godefroit, P. (Ed.). Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press. ss. 467-487.
- ^ Holtz, Thomas R. Jr. (2012) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix. 12 Ağustos 2017 tarihinde Wayback Machine sitesinde arşivlendi.
- ^ a b Lee (2014). "Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus". Nature. 515 (7526): 1-4. doi:10.1038/nature13874. PMID 25337880.
- ^ "Discover | Natural History Museum". www.nhm.ac.uk (İngilizce). 11 Ocak 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Ağustos 2021.
- ^ Choiniere (2012). "New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous Kirkwood Formation in South Africa". Journal of African Earth Sciences. 71–72: 1-17. doi:10.1016/j.jafrearsci.2012.05.005.
- ^ "Last of the Dinosaurs: The Cretaceous Period". 17 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Ağustos 2021.
- ^ van der Reest (2016). "A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada". Cretaceous Research. 58: 108-117. doi:10.1016/j.cretres.2015.10.004.
- ^ Lee (2014). "Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus". Nature. 515 (7526): 257-260. doi:10.1038/nature13874. PMID 25337880.
- ^ Foth (2014). "New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers". Nature. 511 (7507): 79-82. doi:10.1038/nature13467. PMID 24990749.
- ^ Van Der Reest (2016). "A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada". Cretaceous Research. 58: 108-117. doi:10.1016/j.cretres.2015.10.004.
- ^ "Taxon Search: TaxonSearch Archive Page - Stem Archosauria 1.0". web.archive.org. 15 Ocak 2009. 8 Ağustos 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Ağustos 2021.
- ^ Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. 2002.
- ^ Hartman, S.; Mortimer, M.; Wahl, W. R.; Lomax, D. R.; Lippincott, J.; Lovelace, D. M. (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247. doi:10.7717/peerj.7247
 . PMC 6626525 $2. PMID 31333906.
. PMC 6626525 $2. PMID 31333906. - ^ Claudia Inés Serrano-Brañas (2020). "Paraxenisaurus normalensis, a large deinocheirid ornithomimosaur from the Cerro del Pueblo Formation (Upper Cretaceous), Coahuila, Mexico". Journal of South American Earth Sciences. 101: Article 102610. doi:10.1016/j.jsames.2020.102610.
- ^ Nicholls and Russell (1985)
- ^ Schmitz and Motani (2011)
- ^ Gilmore (1933). "On the dinosaurian fauna of the Iren Dabasu Formation". Bulletin of the American Museum of Natural History. 67 (2): 23−78.
- ^ Kobayashi (2003). "A new ornithomimid dinosaur with gregarious habits from the Late Cretaceous of China" (PDF). Acta Palaeontologica Polonica. 48 (2): 235−259. 31 Ağustos 2021 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 17 Ağustos 2021.
- ^ Chinzorig (2017). "Multitaxic bonebed of two new ornithomimids (Theropoda, Ornithomimosauria) from the Upper Cretaceous Bayanshiree Formnation of southeastern Gobi desert, Mongolia". Journal of Vertebrate Paleontology. Program and Abstracts: 97.
- ^ Evolution, diversity, and disparity of ornithomimosaurs (Dinosauria: Theropoda) from the Upper Cretaceous of Mongolia (PDF) (PhD thesis tez). Hokkaido University. 2019. 17 Ekim 2020 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 17 Ağustos 2021.
Konuyla ilgili yayınlar
- Barrett (2005). "The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria)". Palaeontology. 48 (2): 347-358. doi:10.1111/j.1475-4983.2005.00448.x.
- British Museum (Doğa Tarihi): Devekuşu Dinozorları 24 Eylül 2014 tarihinde Wayback Machine sitesinde arşivlendi.
- Jacobsen, AR 2001. Dişle işaretlenmiş küçük theropod kemiği: Son derece nadir bir iz. P. 58-63. İçinde: Mezozoik Omurgalı Yaşamı . Ed.s Tanke, DH, Carpenter, K., Skrepnick, MW Indiana University Press.
- Li Xu (2011). "A new ornithomimid dinosaur with North American affinities from the Late Cretaceous Qiupa Formation in Henan Province of China". Cretaceous Research. 32 (2): 213-222. doi:10.1016/j.cretres.2010.12.004.
- Molnar, RE, 2001, Theropod paleopatology: a literatür araştırması: In: Mesozoic Vertebrate Life, Editör Tanke, DH ve Carpenter, K., Indiana University Press, s. 337-363.
- Nicholls (1985). "Structure and function of the pectoral girdle and forelimb of Struthiomimus altus (Theropoda: Ornithomimidae)". Palaeontology. 28: 643-677.
- Norell (2001). "The beaks of ostrich dinosaurs". Nature. 412 (6850): 873-874. doi:10.1038/35091139. PMID 11528466.
- Schmitz, L. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. 332 (6030): 705-8. doi:10.1126/science.1200043. PMID 21493820.
- Sereno, PC 2005. Stem Archosauria—TaxonSearch [versiyon 1.0, 2005 7 Kasım]
- Tanke, DH ve Brett-Surman, MK 2001. Dinozor İl Parkı'ndan (Dinozor Parkı Oluşumu: Campanian), Alberta, Kanada'dan Kuluçka ve Yuva Boyu Hadrosaurların (Reptilia:Ornithischia) Kanıtı. s. 206-218. İçinde: Mezozoik Omurgalı Yaşamı—Philip J. Currie'nin Paleontolojisinden Esinlenen Yeni Araştırma. DH Tanke ve K. Carpenter tarafından düzenlendi. Indiana University Press: Bloomington. xviii + 577 s.
- Turner (2007). "Supporting online material for: A basal dromaeosaurid and size evolution preceding avian flight". Science. 317 (5843): 1378-1381. doi:10.1126/science.1144066. PMID 17823350. (ek)