Lüsiferin

Lüsiferin (Latince lucifer, "ışık getiren"[1]), biyolüminesans yoluyla ışık yayan canlılarda bulunan ışığın üretilmesinden sorumlu doğal bileşiklere verilen genel bir addır. Lüsiferin, lüsiferaz adlı bir enzimin katalizörlüğünde oksijen ile reaksiyona girer (oksidasyona uğrar) ve uyarılmış hale geçer. Uyarılmış halden temel hal düzeyine geri dönüş esnasında ışık şeklinde enerji salınımı gerçekleşir. Salınan ışığın dalga boyu ve şiddeti lüsiferinin türüne bağlıdır. En çok bilineni ateş böceği lüciferini, D-lüsiferin, olmakla birlikte çeşitli kimyasal yapıdaki çeşitleri mantarlar, bakteriler ve çeşitli deniz canlılarında da bulunur.
Tarihçe

Biyolüminesan ışıma Antik Yunan ve Roma zamanından başlayarak yakın şekilde gözlemlenmiş ve soğuk ışık olarak adlandırılmıştır.[3] Bu soğuk ışık için havanın gerektiği 1667 yılında Robert Boyle tarafından belgelendi, fakat reaksiyonun gerçekleşmesi için oksijen gerektiği o dönemde henüz oksijen keşfedilmediği için anlaşılamadı.[3] 18. yy başlarında René Réaumur kurutulmuş canlı örneklerine su eklediğinde biyolüminesan ışıma elde etmişti. İki molekülün etkileşiminin ışımanın kaynağı olduğu ise ilk kez Raphael Dubois tarafından ortaya atıldı,[3] lüsiferin ve lüsiferaz kelimeleri de bugünkü anlamında ilk kez 19. yüzyılda[1][3] yine onun tarafından kullanıldı. Dubois topladığı örneklerden ışıma için gerektiğini anladığı iki parçayı özütlemişti. Bunlardan sıcaklıktan zarar görmeyeni luciferin, sıcaklıktan zarar göreni luciferase olarak adlandırdı. Dubois bu iki parçayı kullanarak oksijen varlığında biyolüminesansı taklit edebiliyordu.[4] Daha sonrasında lüsiferin-lüsiferaz etkileşimi Edmund Newton Harvey tarafından 20. yy'ın başlarında çalışıldı. Harvey ikili etkileşimin seçici olduğunu, bir türün lüsiferazının başka bir türün lüsiferini ile etkileşime giremediğini keşfetti.[5] İlk kez lüsiferin izolasyonu 1956 yılında ateş böceği lüsiferini için Green ve McElroy tarafından yapıldı.[6] Lüsiferin ve yapısı üzerine diğer bazı ufuk açıcı çalışmalar Nobel Ödüllü Osamu Shimomura tarafından 1950'li yıllarda yapıldı.
Özellikler ve Mekanizma
Karanlıkta ışıma özelliği 800 farklı cinsten yaklaşık 10,000 türde görülmüştür ve bundan fazlası olduğu tahmin edilmektedir, fakat biyolüminesan ışımanın mekanizması evrimsel süreçte korunmamıştır ve farklı türlerdeki lüsiferaz enzimleri DNA dizilimi açısından homolog değildir.[7] Biyolüminesans farklı türler için tekrar ve tekrar evrilmiştir. Filogenik çalışmalar yaklaşık 30 bağımsız şekilde gelişmiş lüsiferin-lüsiferaz sistemi olduğunu ortaya koymaktadır ve bunlardan sadece 9'unun mekanizması üzerinde çalışılmıştır.[8] Bu sistemler tarafından üretilen ışığın dalga boyu 400-700 nm arasında geniş bir açıklığa, yani kırmızıdan mavi renk tonlarına uzanan bir çeşitliliğe sahiptir.[9] Mavi renk tonlarında ışıma kırmızı renk tonlarına göre daha sık karşılaşılmaktadır.[9] Mavi tonların yoğunluğu biyolüminesansın genellikle deniz canlılarında görülmesi ve mavi ışığın su içindeki geçirgenliğinin daha fazla olmasına bağlanabilir.
Işığın üretilmesi lüsiferin molekülünün lüsiferaz enzimi tarafından oksijen ile yükseltgenmesine bağlıdır ve bu reaksiyonun gerçekleşmesi türden türe değişmekle birlikte ATP ve bazı diğer kofaktörlerin varlığını gerektirir. Yükseltgenme sonucunda yüksek enerjili ara ürün oluşur. Kısa süre içinde yüksek enerjili ara ürün temel hale geri döner ve foton üretilir. Reaksiyon sonucunda lüsiferin tüketilir.
![{\textstyle {\ce {Lusiferin + O2 ->[{Lusiferaz}][{ATP, kofaktorler}] oxy-Lusiferin + h\nu}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/88aea7d94961e6b90c1e7c9a74e104469a44ff9c)
Yüksek enerjili ara ürün genellikle dörtlü bir halka olan dioxetane (α-peroxylactone) yapısından dekarboksilasyon ile molekülünün uzaklaştırılması sonucu oluşur. Lüsiferin yapıları değişiklik göstermekle birlikte genellikle dioksietan yapısını içerir.


Lüsiferin türleri
Coelenterazine Bazlı Sistemler


Biyolüminesan ışıma yapan organizmalar arasında çeşitlilik en çok deniz ekosisteminde görülür. Bağımsız şekilde evrimleşmiş pek çok lüsiferaz varyasyonuna rağmen bu organizmalar genellikle aynı lüsiferin varyasyonunu, coelenterazine, kullanırlar.[10] Coelenterazine iki tirosin ve bir fenilalaninden oluşan bir tripeptitin değişmiş halidir.[10] Coelenterazine bazlı sistemi kullanan pek çok tür coelenterazine'i kendi başına sentezlemez ve besin yoluyla alır.[10] Bu sistemle üretilen ışık çoğunlukla 450-500 nm aralığında, mavi renktedir ve genellikle oksijen dışında ek bir kofaktör gerektirmez.[10] Bazı türler biyolüminesan ışığın rengini etkileşime giren ek floresans proteinler kullanarak değiştirebilirler.[10]
Coelenterazine nötral solüsyonda oldukça kararlı olmasına rağmen oksijenle kolayca reaksiyona girer. Oksijen ile reaksiyonunda coelenteramide oluşur. Coelenterazine oksidasyonu enzim yoluyla yapılabildiği gibi (biyolüminesans) kendiliğinden de olabilir (kemilüminesans). Coelenterazine kullanan bazı lüsiferaz tipleri ve özellikleri tablo 1'de görülebilir.
| Tip | Moleküler Ağırlık (kDa) | Organizma | Biyolojik Uygulamaları |
|---|---|---|---|
| Renilla luciferase[11][12][13] | 36 | Bir mercan türü | Biyo-görüntüleme İlaç izleme |
| Gaussia luciferase[14][15] | 20 | Copepoda | İlaç etkisi takibi Tümör büyümesi izleme |
| Nanoluc luciferase[16] | 19 | Oplophorus gracilirostric | Hücre ve doku görüntüleme |
Cypridina Lüsiferin Bazlı Sistem

Cypridina lüsiferin, coelenterazine gibi üç aminoasitin oluşturduğu bir tripeptitin değişmiş halidir. Biyolojik sentez yolu bilinmemekle beraber triptofan, izolösin ve arjinin aminoasitlerinden oluştuğu bilinmektedir.[17] Cypridina (ya da Vargula) lüsiferin bazlı sistemin oluşturduğu ışık mavi renk tonlarına sahiptir.[10] Bir ostrakod olan Cypridina (ya da Vargula) ve Porichthys isimli biyolüminesan balıkta görülür.[10] Vargula hilgendorfii (ya da Cypridina hilgendorfii[18]) bir tehlike sezdiğinde suya lüminesan bir sıvı salgılar. Cypridina bazlı sistem biyogörüntüleme[19] ve bağışıklık testlerinde[20] kullanıldı.
D-lüsiferin Bazlı Sistemler

D-lüsiferin bazlı sistemler en çok çalışılmış biyolüminesan sistemler olmakla birlikte aynı zamanda uygulamada da oldukça kullanışlıdır. Biyolüminesans dört farklı böcek sınıfında görülür: Collembola, Hemiptera, Coleoptera ve Diptera. Coleoptera sınıfından üç farklı aile ışık üretebilir: Phengodidae, Elateridae ve Lampyridae.[21] Amerikan ateş böceği Photinus pyralis, Lampyridae ailesindendir. Ateş böceği üzerindeki ilk biyolüminesan araştırmalar 1917'de Harvey tarafından başladı.

D-lüsiferin kararlı ve toksik olmayan bir moleküldür[22] ve ağırlığı 280.32 g/mol'dür. Eşlenik lüsiferazının ağırlığı 60-62 kDa arasında değişir. Reaksiyon sonucunda sarı, turuncu ve bazı durumlarda kırmızı ışık üretir. Bu durum biyolüminesan sistemlerin paralel evriminin başka bir göstergesidir.[23] Suda yaşayan canlılarda mavi ışığın evrilmesinin sebebi mavi ışığın suda daha uzaktan görülebilmesi olduğu gibi, kırmızıya yakın tonların kara ortamında evrilmesi de uzun dalga boylu ışığın hava ortamında daha uzaktan görülebilmesidir.
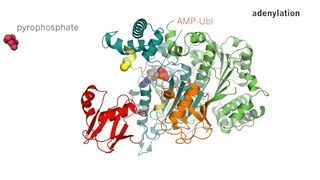
Lüsiferaz enzimiyle olan reaksiyonunda D-lüsiferin'in oksijene ek kofaktör olarak ve ATP'ye ihtiyaç duyduğu 1940'larda William D. McElroy tarafından gösterildi.[21] Bu reaksiyonun son ürünleri AMP, iki serbest fosfat, karbondioksit ve ışıktır.

![{\displaystyle {\ce {LH2 + O2 + ATP ->[{Lüsiferaz}][Mg^+2] oxy-L +CO2 +AMP + PPi + h\nu}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b137da273e206a29839bb2ab115f579680f84283)
Reaksiyonda üretilen ışık gerçekleştiği canlı türüne göre 530 nm ile 635 nm arasındadır. Bununla birlikte değişik pH'larda uyarımı sonucu floresan yayınımı 537 nm olarak ölçülmüştür.[24] Hücre içinde pH 7.8 ve 23-25 °C de gerçekleşen reaksiyon laboratuvar ortamında daha geniş bir aralıkta çalışır ve asidik ortamda 615 nm boyunda ışık üretmiştir.


-
Tetrapyrrole Bazlı Lüsiferinler
Bakteriyel Biyolüminesan Sistem
Mantarsal Biyolüminesan Sistem
Türler
Lusiferinler, bir lusiferaz (bir enzim) varlığında oksijenle reaksiyona girerek enerji'yi ışık şeklinde serbest bırakan küçük-moleküllü substratlar sınıfıdır. Kaç tane lüsiferin türü olduğu bilinmemektedir ama daha iyi çalışılmış bileşiklerden bazıları aşağıda listelenmiştir.
Lusiferinlerin kimyasal çeşitliliği nedeniyle, hepsinin moleküler oksijen gerektirmesi dışında net bir birleştirici etki mekanizması yoktur.[25] Lusiferinlerin ve lusiferazların çeşitliliği, çeşitli reaksiyon mekanizmaları ve dağınık filogenetik dağılımı, birçoğunun bağımsız olarak evrim sürecinde ortaya çıktığını gösterir.[25]
Ateşböceği
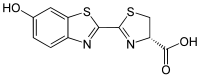
Ateşböceği lusiferini, birçok Lampyridae türünde bulunan lusiferindir.
Böcek lusiferazları (EC 1.13.12.7)'nin substratıdır ve ateşböceklerinden gelen karakteristik sarı ışık emisyonundan sorumludur ama diğer yandan ışık yaymayan türlerden ilgili enzimlerle ışık üretmek için çapraz reaksiyona girebilir.[26] Işık emisyonu için moleküler oksijen'e ek olarak adenosin trifosfat (ATP) gerektiğinden, kimyası olağandışıdır.[27]
Uygulamalar
Literatür
Kaynakça
- ^ a b “Luciferin.” The Merriam-Webster.com Dictionary, Merriam-Webster Inc., https://www.merriam-webster.com/dictionary/luciferin. Accessed 15 January 2020.
- ^ Harvey, E.N. (1957). A history of luminescence from the earliest times until 1900. vol 44. The American Philosophical Society, Philadelphia.
- ^ a b c d Lee, J. (2014). A History of Bioluminesence, University of Georgia. Avaliable at: http://photobiology.info/HistBiolum/HistBiolum.html 8 Ocak 2020 tarihinde Wayback Machine sitesinde arşivlendi. Accessed at: 16/01/2020
- ^ Waldemar Adam: Biologisches Licht. In: Chemie in unserer ZEit. Band 7, Nr: 6, 1973. S. 182-192, doi:10.1002/ciuz.19730070605.
- ^ Harvey: Bioluminescence. Academic, New York 1952.
- ^ Green A.A. and W.D. McElroy, W.D. (1956) Biochim. Biophys. Acta 20: 170-176.
- ^ Haddock SHD, Moline MA, Case JF (2010) Bioluminescence in the sea. Annu Rev Mar Sci 2:443–493
- ^ Kaskova ZM, Tsarkova AS, Yampolsky IV (2016) 1001 Lights: lucif-erins, luciferases, their mechanisms of action and applications in chemical analysis, biology and medicine. Chem Soc Rev. https ://doi.org/10.1039/c6cs0 0296j
- ^ a b Eveline Brodl, Andreas Winkler, Peter Macheroux: Molecular Mechanisms of Bacterial Bioluminescence. In: Computational and Structural Biotechnology Journal. Band 16, 15. November 2018, S. 551-564 doi:10.1016/j.csbj.2018.11.003
- ^ a b c d e f g Fleiss, A., Sarkisyan, K.S. A brief review of bioluminescent systems (2019). Curr Genet 65, 877–882 (2019) doi:10.1007/s00294-019-00951-5
- ^ Lorenz WW, McCann RO, Longiaru M, Cormier MJ (1991) Isolation and Expression of a cDNA Encoding Renilla Reniformis Luciferase. Proc Natl Acad Sci USA 88(10):4438–4442
- ^ Hamdan FF, Audet M, Garneau P, Pelletier J, Bouvier M (2005) High-throughput screening of G protein-coupled receptor antagonists using a bioluminescence resonance energy transfer 1-based β-arrestin2 recruitment assay. J Biomol Screen 10(5):463–475
- ^ Prinz A, Diskar M, Herberg FW (2006) Application of biolumines-cence resonance energy transfer (BRET) for biomolecular inter-action studies. Chembiochem Eur J Chem Biol 7(7):1007–1012
- ^ Tannous BA, Kim DE, Fernandez JL, Weissleder R, Breakefield XO (2005) Codon-optimized Gaussia luciferase cDNA for mammalian gene expression in culture and in vivo. Mol Therapy J Am Soc Gene Therapy 11(3):435–443
- ^ Chung E, Yamashita H, Au P, Tannous BA, Fukumura D, Jain RK (2009) Secreted Gaussia luciferase as a biomarker for monitoring tumor progression and treatment response of systemic metastases. PloS One 4(12):e8316
- ^ Saito K, Chang YF, Horikawa K, Hatsugai N, Higuchi Y, Hashida M, Yoshida Y, Matsuda T, Arai Y, Nagai T (2012) Luminescent proteins for high-speed single-cell and whole-body imaging. Nature Communications 3:1262
- ^ Oba Y, Kato SI, Ojika M, Inouye S (2002) Biosynthesis of luciferin in the sea firefly, cypridina hilgendorfii: l-tryptophan is a component in cypridina luciferin. Tetrahedron Lett 43(13):2389–2392
- ^ J. G. Morin: Based on a review of the data, use of the term 'cypridinid' solves the Cypridina/Vargula dilemma for naming the constituents of the luminescent system of ostracods in the family Cypridinidae. In: Luminescence. 26(1), 2011, S. 1–4. PMID 19862683; doi:10.1002/bio.1178
- ^ Yamada Y, Nishide SY, Nakajima Y, Watanabe T, Ohmiya Y, Honma KI, Honma S (2013) Monitoring Circadian time in rat plasma using a secreted cypridina luciferase reporter. Anal Biochem 439(2):80–87
- ^ Wu C, Irie S, Yamamoto S, Ohmiya Y (2009) A Bioluminescent enzyme immunoassay for prostaglandin E(2) using cypridina luciferase. Lumin J Biol Chem Lumin 24(2):131–133
- ^ a b Osamu Shimomura, S. 1ff.
- ^ Tiffen JC, Bailey CG, Ng C, Rasko JEJ, Holst J (2010) Luciferase expression and bioluminescence does not affect tumor cell growth in vitro or in vivo. Mol Cancer 9:299
- ^ Fallon TR, Lower SE, Chang CH, Bessho-Uehara M, Martin GJ, Bewick AJ, Behringer M et al (2018) Firefly genomes illuminate parallel origins of bioluminescence in beetles. eLife. https://doi.org/10.7554/eLife.36495
- ^ Bowie, L.J., Synthesis of Firefly Luciferin and Structural Analogs. Methods in Enzymology, 57, 23 (1978).
- ^ a b Hastings JW (1983). "Biological diversity, chemical mechanisms, and the evolutionary origins of bioluminescent systems". Journal of Molecular Evolution. 19 (5): 309-321. Bibcode:1983JMolE..19..309H. doi:10.1007/BF02101634. PMID 6358519.
- ^ Viviani VR, Bechara EJ (1996). "Larval Tenebrio molitor (Coleoptera: Tenebrionidae) Fat Body Extracts Catalyze Firefly D-Luciferin-and ATP-Dependent Chemiluminescence: A Luciferase-like Enzyme". Photochemistry and Photobiology. 63 (6): 713-718. doi:10.1111/j.1751-1097.1996.tb09620.x.
- ^ Green A, Mcelroy WD (October 1956). "Function of adenosine triphosphate in the activation of luciferin". Archives of Biochemistry and Biophysics. 64 (2): 257-271. doi:10.1016/0003-9861(56)90268-5. PMID 13363432.