Kaspari şeridi
Kaspari şeridi, vasküler bitkilerin ( Pteridophytes[1] ve Spermatophytes ) kök endodermisinin (radyal ve hücre duvarları ) merkezinde bant benzeri bir kalınlaşmadır. Bölgenin bileşimi esas olarak lignindir ve genişliği türler arasında değişmektedir. Kaspariişeridi su geçirmezdir, bu nedenle korteks ve damar demeti arasında su ve inorganik tuzların taşınmasını kontrol edebilir, su ve inorganik tuzların apoplast yoluyla stele taşınmasını önler, böylece hücre zarına girip hareket etmesi gerekir. Simplastik yoldan stele hücrenin iç ve dış nesnelerini bloke eder.[] Toplu taşımanın işlevi hayvan dokularınınkine benzer.[] .[2][3] Kaspari şeridinin gelişimi, SHORT-ROOT (SHR), SCARECROW (SCR) ve MYB36 gibi transkripsiyon faktörlerinin yanı sıra orta kolon hücreleri tarafından sentezlenen polipeptit hormonu tarafından düzenlenir.[4][5]


Kaspari şeridinin kimyası, süberinden oluşan kısım tarif edilmiştir. Bazı araştırmalara göre,[6] Kaspari şeridi, kısmen oksitlenmiş filmler olarak radyal duvarlar arasındaki orta lamelde fenolik ve doymamış yağlı maddelerin lokalize bir birikimi olarak oluşmaya başlar. Birincil duvar, bu duvarın iç kısmındaki benzer maddelerin tortularıyla kaplanır ve daha sonra kalınlaşır. Hücre duvarının Hazar şeridini oluşturan malzeme tarafından kaplanması bu yol boyunca kılcal hareket yoluyla su ve besinlerin hareketine izin verecek olan gözenekleri tıkar.[7] Endodermal hücrenin sitoplazması, hücreler protoplastların büzülmesine maruz kaldığında şeritten kolayca ayrılmaması için kaspari şeridine sıkıca bağlıdır. Kökte, kaspari şeridi, kök ucunun arkasındaki kökün büyümeyen bölgesindeki endodermal hücrelerin hücre duvarı içine gömülür.[8] Burada, kaspari şeridi, korteksin apoplastını vasküler dokunun apoplastından ayıran ve böylece ikisi arasındaki materyalin difüzyonunu bloke eden bir sınır tabakası görevi görür.[9] Bu ayırma, endodermis tabakasını geçmek için suyu ve çözünenleri simplastik yoldan plazma zarından geçmeye zorlar.[8]
Korteksin dışa doğru büyümesi tamamlandıktan sonra kaspari şeritleri farklılaşır. Kök gelişiminin bu seviyesinde, vasküler silindirinin birincil ksilemi sadece kısmen ilerlemiştir. Gymnospermlerde ve sekonder büyüme gösteren anjiyospermlerde, kökler genellikle sadece kaspari şeritleri bulunduran endodermis oluşturur. Bunların birçoğunda, endodermis daha sonra perisiklden periderm geliştiğinde korteksle birlikte atılır. Perisikl yüzeysel ise ve korteks tutulursa, ya endodermis gerilir veya ezilir ya da radyal antiklinal bölünmelerle vasküler silindirin genişlemesine ayak uydurur ve yeni duvarlar eskileriyle süreklilik içinde kaspari şeritleri geliştirir.[10]
İkincil büyümenin yokluğunda (çoğu monokotiledon ve birkaç eudicot), endodermis genellikle duvar modifikasyonlarına uğrar. Kaspari şeridinin gelişiminin ötesinde iki gelişim aşaması vardır. İkinci aşamada suberin (veya endoderm[7] ) hücrenin içindeki tüm duvarı kaplar. Sonuç olarak, kaspari şeridi sitoplazmadan ayrılır ve ikisi arasındaki bağlantı belirgin olmaktan çıkar. Üçüncü aşamada, suberin üzerinde, bazen esas olarak iç teğet duvarlarda kalın bir selüloz tabakası biriktirilir. Kalınlaştırılmış duvar ve Hazar şeridinin bulunduğu orijinal duvar odunlaşarak ikincil bir hücre duvarı oluşturabilir. Kaspari şeridi, endodermal duvarın kalınlaşması meydana geldikten sonra tanımlanabilir. Kalınlaşmış endodermal duvarda çukurlar olabilir. Endodermal duvarların ardışık gelişimi, monokotiledonlarda açıkça ifade edilir.[11][12]
Keşif
Kaspari şeridinin keşfi, 19. yüzyılın ortalarına kadar uzanır ve bitkilerin kökünün endodermisinin anlaşılmasında ilerlemeler sağlar.[13] 1865'te Alman botanikçi Robert Caspary, bitki kökünün endodermisini ilk kez tanımladı, hücre duvarının kalınlaştığını keşfetti ve ona Schuchtzscheide adını verdi. Daha sonra bilim adamları, Casbury'nin adını taşıyan kalınlaştırılmış kısmına Carls Kuşağı adını verdiler.[3][14] "Caspary'schen fleck" terimi ( German ) 1870'lerin literatüründe[15][16] ortaya çıktı ve 20. yüzyıldan sonra genellikle Hazar şeridi olarak adlandırıldı. 1922'de araştırmacılar, kompozisyonunu incelemek için ilk olarak bitkilerin kökünden Hazar şeridini terk ettiler.[][17][18]
Kompozisyon
Kaspari şeridinin kimyasal bileşimi uzun süredir tartışmalıdır. Casbury, bu yapının lignin veya süberinden oluşabileceğine dikkat çekti. Daha sonraki bilim adamları çoğunlukla bunun süberin olduğunu düşündüler.[19] Birkaç bitkinin Hazar şeridini analiz ettikten sonra, ligninin ana bileşen olduğu 1990'lara kadar değildi, ancak birçok ders kitabı güncellenmedi.[2] Endotelin hücre duvarı odunsu doku açısından zengin olmasına rağmen, bu endotelin alt düzey farklılaşmasının sonucudur. Geçmişte, bazı bilim adamları, Hazar şeridinin endodermisinin oluşumunun alt düzey farklılaşmasının başlangıcı olduğuna inanırlar ancak ikisi arasında doğrudan bir ilişki yoktur. Kaspari şeridinin birincil farklılaşmasından sonra oluşmuştur ve ikincil farklılaşma, kaspari şeridinin olduğu yerde değil, kökün eğik çizgiyle kesilmesiyle başlar.[1]
İşlev
Kaspari şerit, orta lamel de dahil olmak üzere endotel hücreleri arasındaki boşlukla tamamen doldurulur ve iki hücrenin hücre duvarlarını neredeyse kaynaştırır.[1] Bitkilerin kökünde su ve inorganik besinlerin taşınmasında, kaspari şeridi esas olarak birincil in vitro taşınmasını, yani suyun ve inorganik tuzların epidermis ve korteks hücrelerinin interstisyel hücreleri yoluyla taşınmasını etkiler. Endotel hücrelerine su ve inorganik tuz geldiğinde, kaspari şeridi su geçirgen olmadığı için hücre zarından hücreye girmeleri gerekir ve daha sonra stelin lignan hücrelerine ulaşmak için protoplazmik iç yol ile taşınır ve daha sonra saplar ve yapraklar gibi diğer organlara taşınır.[14] Büyüme ortamı elverişsiz olduğunda, kaspari şeridi, bitki hücreleri ile dış dünya arasında bir bariyer görevi görerek, çevreye iyonların girmesini veya kendi iyonlarının dışarı çıkmasını önleyebilir.[5] Ayrıca karkas kuşağının ve korteksin kalınlaşması, su dağılımını önleme işlevinin yanı sıra toksik madde veya patojen istilasını da engeller.[20] Bazı çalışmalar, bitkilerin yüksek tuzlu ortamlarda ve çevreye bir adaptasyon olabilecek köklerin ucuna daha yakın alanlarda,[21][22] ancak farklılaşmış endotelyal alt seviye ile karşılaştırıldığında, daha kalın kaspari şeridi oluşturabileceğini göstermiştir. Yüksek tuz sıkıntısında önemli ölçüde kalınlaşan odunsu duvarlarda, kaspari şeridi farklılaşmaları daha küçüktür.[23]
Kaspari şeridi esas olarak kökün ekzodermisinde bulunur,[24] ancak bazı bitkilerde ayrıca kök korteksinin, gövdenin veya yaprağın dış tarafında dış kortekste kaspari şeridi bulunur.[25] Örneğin, Pinus bungeana'nın kozalaklı ağaçları ve Pelargonium'un gövdeleri, su dağılımını veya patojenik istilayı önleme ile ilgili olabilecek kaspari şeridine sahiptir.[26][27]
Gelişim

Carters kayışının gelişimi, endojenik hücreler tamamen geciktirildikten sonra başlar[19][28] ve şu anda kaspari şeridinin endotelyal hücre oluşumunu destekleyen iki haber sinyal iletimi vardır. Birincisi transkripsiyon faktörü Kısa kök (SHR) Aktive edilmiş ek iki transkripsiyon faktörü Myb36 ve Korkuluk (SCR), birincisi kaspari şeridini Strip Proteinlerini (Casp1-5), Peroksidaz (PER64) ve ESB1 (Gelişmiş) Suberin 1), vb. uyarabilir ., ikincisi, ikincisi, iç deri hücresindeki karbür kayışın konumunu etkiler, bu da kaspari şeridinin konumunun Stele çok yakın olmasına neden olur;[4] ikincisi orta Casparian Strip Integrity Factor (CIF1-2) ve endotel hücre radyal duvarına bağlı GSO1 (SGN3) ve GSO2 reseptörü ve yan duvardaki GSO2 reseptörüdür. Hücrelerdeki CASP, Cascade Band'ın konumuna karşılık gelen bir hücre membran bölgesine konsantre edilir ve bir Casparian Strip Membran Domain (CSD) oluşturur ve CSD bölgeye dahil edilir, GS01 reseptörü her CSD'nin kenarı ile çevrilidir. sürekli bir şerit bölgesine kaynaşmış CSD'yi teşvik eden bölge, yani kaspari şeridinin oluşturulacağı bölge.[5][29]
Kaspari şeridi proteini, birbiriyle etkileşime giren ve PER64, ESB1 ve solunum oksidaz homologeri F (RBOHF) gibi lignini sentezlemek için gerekli proteinlere bağlanarak Karzit bant gelişiminin akış aşağı reaksiyonunu aktive edebilen bir zar proteinidir .[1][3] GSO1 reseptörlerinden yoksun veya aynı zamanda CIF1 ve CIF2 polipeptitlerinden yoksun mutant bitkilerde, CASP1 endotel hücre zarı üzerinde anormal bir şekilde dağılmıştır ve CSD normal olarak sürekli ve tam bir bant yapısı oluşturamaz, bu nedenle sonunda kırık ve süreksiz bir Carlsian bandı oluşturur.[5][29]
Işık, toprak tuzluluğu ve su eksikliği gibi çevresel faktörler, kaspari şeridinin gelişimini etkileyebilir.[26]
Fotoğraf
 Acorus cinsinin ( monokotiledonlar ) kökünün damar demetleri endotelde ve orta kolonun etrafındaki Casparian şeridinde görülebilir. Kasparian şeridi, lignin içerdiği için kırmızıya boyanmıştır.
Acorus cinsinin ( monokotiledonlar ) kökünün damar demetleri endotelde ve orta kolonun etrafındaki Casparian şeridinde görülebilir. Kasparian şeridi, lignin içerdiği için kırmızıya boyanmıştır. Ranunculus ( dicotyledon ) cinsinin kökünün damar demeti endotelini ve orta kolonun etrafındaki Casparian şeridini görebilir. Kasparian şeridi, lignin içerdiği için kırmızıya boyanmıştır.
Ranunculus ( dicotyledon ) cinsinin kökünün damar demeti endotelini ve orta kolonun etrafındaki Casparian şeridini görebilir. Kasparian şeridi, lignin içerdiği için kırmızıya boyanmıştır.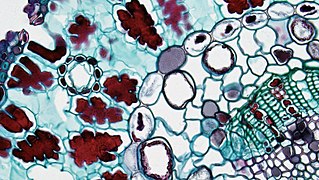 Pinaceae ( Gymnosperm ) iğne yapraklı yapraklar. Periferik endotelyal endodermis ve görünür vasküler demetlerden oluşan Casparian şeridi
Pinaceae ( Gymnosperm ) iğne yapraklı yapraklar. Periferik endotelyal endodermis ve görünür vasküler demetlerden oluşan Casparian şeridi
Ayrıca bakınız
- süberin
- endodermis
- ekzodermis
Notlar
Şablon:Notefoot
Kaynakça
- ^ a b c d Geldner (2013). "The Endodermis". Annual Review of Plant Biology. 64 (1): 531-558. doi:10.1146/annurev-arplant-050312-120050. ISSN 1543-5008. PMID 23451777.
- ^ a b Geldner, N. (2013). "Casparian strips" (PDF). Current Biology. 23 (23). ss. R1025, R1026.
- ^ a b c Roppolo (2011). "A novel protein family mediates Casparian strip formation in the endodermis". Nature. 473 (7347): 380-383. doi:10.1038/nature10070. ISSN 0028-0836. PMID 21593871.
- ^ a b Li, P. (2018). "Construction of a Functional Casparian Strip in Non-endodermal Lineages Is Orchestrated by Two Parallel Signaling Systems in Arabidopsis thaliana". Current Biology. 28 (17): 2777-2786.e2. doi:10.1016/j.cub.2018.07.028. PMID 30057307.
- ^ a b c d Nakayama, T. (2017). "A peptide hormone required for Casparian strip diffusion barrier formation in Arabidopsis roots". Science. 355 (6322): 284-286. doi:10.1126/science.aai9057. PMID 28104889.
- ^ Van Fleet (1961). "Histochemistry and function of the endodermis". Botanical Review. 27 (2): 165-220. doi:10.1007/BF02860082.
- ^ a b Cytology of the Ray Cells in Sapwood and Heartwood. Cram. 1959.
- ^ a b Taiz, L., Zeiger, Eduardo, Møller, Ian Max, & Murphy, Angus. (2015). Plant physiology and development (Sixth ed.).
- ^ Grebe (2011). "Unveiling the Casparian strip". Nature. 473 (7347): 294-5. doi:10.1038/473294a. PMID 21593860.
- ^ Die physiologischen Scheiden. Borntraeger. 1943.
- ^ OGURA (1938). "Problems in morphology (13)". Botany and Zoology. 6: 139-148.
- ^ 100 Jahre Köln-düsseldorfer Rheindampfschiffahrt: Insbesondere Zerstörung und Wiederaufbau 1939-1953. Köln-Düsseldorfer Rheindampfshiffahrt. 1953.
- ^ Seago, J. L., Jr. (2020). "Revisiting the occurrence and evidence of endodermis in angiosperm shoots". Flora. 273: 151709. doi:10.1016/j.flora.2020.151709.
- ^ a b Grebe (2011). "Unveiling the Casparian strip". Nature. 473 (7347): 294-295. doi:10.1038/473294a. ISSN 0028-0836. PMID 21593860.
- ^ "Rhizocarpeae. I. Axenorgane: A. Stamm: a. Marsilia (Drummondii, elata, salvatrix)". Vergleichende Untersuchungen betreffend die Histiologie (Histiographie und Histiogenie) der vegetativen und Sporen-bildenden Organe und die Entwickelung der Sporen der Leitbündel-Kryptogamen: mit Berücksichtigung der Histiologie der Phanerogamen, ausgehend von der Betrachtung der Marsiliaceen. Commissionnaires de l'Académie Impériale des sciences. 1872. ss. 1-12.
- ^ Müller, C. (1884). "Morphologie der Gewebe". 12 (1): 234-342. 10 Mayıs 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Ağustos 2022.
- ^ Priestley, J. H.; North, E. E. (1922). "Physiological Studies in Plant Anatomy III. The Structure of the Endodermis in Relation to its Function". The New Phytologist. 21 (3): 113-139. doi:10.1111/j.1469-8137.1922.tb07593.x. 18 Kasım 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Ağustos 2022.
- ^ Song (2019). "Development and chemical characterization of Casparian strips in the roots of Chinese fir (Cunninghamia lanceolata)". Trees. 33 (3): 827-836. doi:10.1007/s00468-019-01820-x. ISSN 0931-1890.
- ^ a b Naseer, S.; Lee, Y.; Lapierre, C.; Franke, R.; Nawrath, C.; Geldner, N. (2012). "Casparian strip diffusion barrier in Arabidopsis is made of a lignin polymer without suberin". Proceedings of the National Academy of Sciences. 109 (25): 10101-6. doi:10.1073/pnas.1205726109. PMC 3382560 $2. PMID 22665765.
- ^ Robbins, N. E.; Trontin, C.; Duan, L.; Dinneny, J. R. (2014). "Beyond the barrier: communication in the root through the endodermis". Plant Physiology. 166 (2): 551-9. doi:10.1104/pp.114.244871. PMC 4213087 $2. PMID 25125504.
- ^ Karahara, I.; Ikeda, A.; Kondo, T.; Uetake, Y. (2004). "Development of the Casparian strip in primary roots of maize under salt stress". Planta. 219 (1): 41-7. doi:10.1007/s00425-004-1208-7. PMID 14986139.
- ^ Chen, T.; Cai, X.; Wu, X.; Karahara, I.; Schreiber, L.; Lin, J. (2011). "Casparian strip development and its potential function in salt tolerance". Plant Signaling & Behavior. 6 (10): 1499-502. doi:10.4161/psb.6.10.17054. PMC 3256377 $2. PMID 21904117.
- ^ Palmgren, M. (2018). "Plant epithelia: What is the role of the mortar in the wall?". PLOS Biology. 16 (12): e3000073. doi:10.1371/journal.pbio.3000073. PMC 6296743 $2. PMID 30517104.
- ^ Evert, R. F. (28 Ağustos 2006). Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development. John Wiley & Sons. s. 6. ISBN 978-0-470-04737-8. 1 Aralık 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Nisan 2021.
- ^ Lersten (1997). "Occurrence of endodermis with a casparian strip in stem and leaf". The Botanical Review. 63 (3): 265-272. doi:10.1007/BF02857952. ISSN 0006-8101.
- ^ a b Wu (2005). "Casparian Strips in Needles are More Solute Permeable than Endodermal Transport Barriers in Roots of Pinus bungeana". Plant and Cell Physiology. 46 (11): 1799-1808. doi:10.1093/pcp/pci194. ISSN 1471-9053. PMID 16170202.
- ^ Meyer, C. J.; Peterson, C. A. (2011). "Casparian bands occur in the periderm of Pelargonium hortorum stem and root". Annals of Botany. 107 (4): 591-8. doi:10.1093/aob/mcq267. PMC 3064534 $2. PMID 21239408.
- ^ Barberon, M. (2017). "The endodermis as a checkpoint for nutrients". New Phytologist. 213 (4): 1604-1610. doi:10.1111/nph.14140. PMID 27551946.
- ^ a b Pfister, A.; Barberon, M.; Alassimone, J.; Kalmbach, L.; Lee, Y.; Vermeer, J. E. M. (2014). "A receptor-like kinase mutant with absent endodermal diffusion barrier displays selective nutrient homeostasis defects". eLife. 3: e03115. doi:10.7554/eLife.03115. PMC 4164916 $2. PMID 25233277.
- Plant Anatomy. John Wiley & Sons. 1965. s. 767. ISBN 9780471244554. Yazar
|ad1=eksik|soyadı1=(yardım)