Homo habilis
| Homo habilis | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
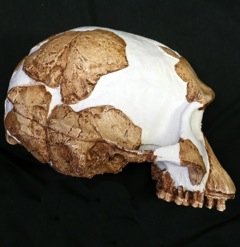
 KNM-ER 1813 adlı örneğin rekonstrüksiyonu, Senckenberg Müzesi, Almanya. | |||||||||||||||||||||||
| Biyolojik sınıflandırma | |||||||||||||||||||||||
| |||||||||||||||||||||||
| İkili adlandırma | |||||||||||||||||||||||
| †Homo habilis Leakey vd., 1964 | |||||||||||||||||||||||
| Sinonimler [1] | |||||||||||||||||||||||
| |||||||||||||||||||||||
Homo habilis ("becerikli insan"), yaklaşık 2.3 ila 1.65 milyon yıl önce Doğu ve Güney Afrika'nın Erken Pleistosen'inden soyu tükenmiş bir arkaik insan türüdür. 1964'te türün tanımlaması üzerine, H. habilis oldukça tartışmalıydı ve birçok araştırmacı, o zamanlar bilinen diğer tek erken hominin olan Australopithecus africanus ile sinonim (eşdeğer) olmasını önerdi; ancak H. habilis, zaman geçtikçe ve ilişkili daha çok keşif yapıldıkça daha çok kabul gördü. 1980'lere gelindiğinde, H. habilis'in bir insan atası olduğu ve doğrudan modern insanlara yol açan Homo erectus'a evrimleştiği öne sürüldü. Bu bakış açısı şimdi tartışılıyor. Güvensiz tür tanımlamasına sahip birkaç örnek H. habilis'e atanmış, bu da türün bölünmesi için argümanlara, yani yalnızca ilkinin geniş destek aldığı "H. rudolfensis" ve "H. gautengensis" adlı yeni türlerin önerilmesine neden olmuştur.
Kendisi ile çağdaş Homo türleri gibi, H. habilis'in beyin büyüklüğü genellikle 500–900 cm3 arasında değişiyordu.[2] H. habilis'in vücut oranları, yalnızca oldukça parçalı iki iskeletten bilinmektedir ve büyük ölçüde daha önceki australopitesinlere (iki ayaklı insansılar) benzer bir anatominin öngörülmesine dayanmaktadır. Bu nedenle H. habilis'in Australopithecus cinsine Australopithecus habilis olarak taşınması da önerilmiştir. Bununla birlikte, H. habilis'in verimsiz uzun mesafe yolculuk yetenekleri olan küçük yapılı bir insan olarak yorumlanmasına meydan okunmuştur. Varsayılan dişi örnek OH 62, geleneksel olarak australopitesinlere benzer oranlar varsayılarak 100–120 cm boyunda ve 20–37 kg ağırlığında olduğu şeklinde yorumlanır ancak modern insan benzeri oranlarda olduğu varsayıldığında, yaklaşık 148 cm ve 35 kg olacağı varsayılır. Bununla birlikte, H. habilis, en azından kısmen, australopitesinler için varsayıldığı gibi ağaçta yaşıyor olabilir. Göreceli erkek ve dişi boyutu kesin olarak bilinmemekle birlikte, erken homininler tipik olarak kalın kıllara ve dişilerden çok daha büyük erkeklerle, belirgin eşeysel dimorfizme sahip olarak yeniden yapılandırılmıştır.
H. habilis, Oldowan taş alet endüstrisini üretti ve esas olarak kasaplık aletlerini kullandı. Australopitesinler ile karşılaştırıldığında, erken Homo'nun genellikle yüksek miktarlarda et tükettiği ve özellikle H. habilis'in leşlerle beslendiği düşünülmektedir. Tipik olarak, erken homininler çok eşli toplumlarda yaşamış olarak yorumlanır ancak bu oldukça kurgusaldır. H. habilis toplumunun günümüz savana şempanzeleri ve babunlarınkine benzer olduğunu varsayarsak, grupların büyük kediler, sırtlanlar ve timsahlar gibi açık savan yırtıcılarına karşı savunmak için birden fazla erkekle birlikte 70-85 üyeli olabilir. H. habilis, H. rudolfensis, H. ergaster /H. erectus ve Paranthropus boisei ile aynı dönemde yaşamıştır.
Taksonomi
Araştırma tarihi

Tanınan ilk kalıntılar - OH 7, 1,75 milyon yıl öncesine tarihlenen kısmi çocuk kafatası, el ve ayak kemikleri - 1960 yılında Tanzanya'nın Olduvai Gorge kentinde Jonathan Leakey tarafından keşfedildi. Bununla birlikte, gerçek ilk kalıntılar - OH 4, bir azı dişi - 1959'da Louis ve Mary Leakey'in (Jonathan'ın ebeveynleri) kıdemli asistanı Heselon Mukiri tarafından keşfedildi, ancak bu o sırada fark edilmedi.[3] Bu zamana kadar, Louis ve Mary 29 yıllarını Olduvai Gorge'da erken hominin kalıntıları için kazı yaparak geçirdiler, ancak bunun yerine Oldowan taş alet endüstrisinin yanı sıra esas olarak diğer hayvan kalıntılarını da buldular. Endüstri,1959'da Paranthropus boisei'ye (döneminin ismiyle "Zinjanthropus") bölgede bulunan ilk ve tek hominin olduğu için atfedildi, ancak bu, OH 7'nin keşfi üzerine revize edildi.[3] 1964'te Louis, Güney Afrikalı paleoantropolog Phillip V. Tobias ve İngiliz primatolog John R. Napier, Avustralyalı antropolog Raymond Dart'ın tavsiyesi üzerine kalıntıları resmen Homo cinsine –Homo habilis ismiyle– atadı. H. habilis, Latince'de yetenekli, kullanışlı, zihinsel olarak becerikli, güçlü anlamına gelir.[4] Numunenin Oldowan ile ilişkisi (daha sonra gelişmiş bilişsel yeteneğin kanıtı olarak kabul edildi), onu Homo olarak sınıflandırmak için gerekçe olarak kullanıldı.[5] OH 7, türün holotip numunesi olarak belirlendi.[4]
Tanımlamadan sonra, H. habilis'in kalıntılarının Australopithecus africanus'a ait olarak (o sırada bilinen diğer tek erken hominin) yeniden sınıflandırılıp sınıflandırılmayacağı, kısmen kalıntıların çok eski olması ve Homo'nun Asya'da evrimleştiği tahmin edildiğinden hararetle tartışıldı. Ayrıca beyin boyutu, 1955'te Wilfrid Le Gros Clark'ın türün Homo cinsi içerisinde değerlendirdiği zamanki boyut önermesinden daha küçüktü.[3][6] H. habilis sınıflandırması, daha fazla fosil örneği ve tür ortaya çıktıkça daha geniş kabul görmeye başladı.[3] 1983'te Tobias, A. africanus'un Paranthropus ve Homo'nun (ikisi kardeş taksonlardı) doğrudan bir atası olduğunu ve A. africanus'un (aşağıda görülebileceği gibi) modern insanlara evrimleşmiş H. erectus'a evrimleşen H. habilis'e evrimleştiğini öne sürdü (bir kladogenez süreci ile). Ayrıca A. africanus ve H. habilis arasında büyük bir evrimsel sıçrama olduğunu ve bunun üzerine insan evriminin yavaş yavaş ilerlediğini, çünkü H. habilis'in beyin boyutunun australopitesin öncüllerine kıyasla neredeyse iki katına çıktığını söyledi.[7]





Birçoğu Tobias'ın modelini kabul etmiş ve Geç Pliyosen'den Erken Pleistosen hominin kalıntılarını Paranthropus ve H. erectus aralığının dışında H. habilis'e atamıştı. Kafatası olmayan elemanlar için bu, net tanısal özellikler olmadığı için boyut bazında yapıldı.[8] Bu uygulamalar nedeniyle, türler için varyasyon aralığı oldukça genişledi ve H. habilis sensu stricto ("dar anlamda") ve H. habilis sensu lato ("geniş anlamda") terimleri kullanılmaya başlandı. Bunu çözmek için 1985 yılında İngiliz paleoantropolog Bernard Wood, 1972'de Kenya'nın Turkana Gölü'nde bulunan ve H. habilis'e atanan nispeten büyük kafatası KNM-ER 1470'in aslında şimdi Homo rudolfensis olarak adlandırılan farklı bir türü temsil ettiğini öne sürdü.[9] Bunun yerine erkek bir örneği temsil ettiği, diğer H. habilis örneklerinin ise dişi olduğu iddia edilmektedir.[10] Güney Afrika'daki Erken Homo örnekleri, çeşitli şekillerde H. habilis veya H. ergaster / H. erectus'a atanmıştır, ancak tür tanımı büyük ölçüde belirsizdir. 2010 yılında, Avustralyalı arkeolog Darren Curoe, Güney Afrikalı erken Homo'yu yeni bir tür olan "Homo gautengensis"e bölmeyi önerdi.[11]
1986'da, parça parça bir iskelet olan OH 62, Amerikalı antropolog Tim D. White tarafından H. habilis kafatası parçalarıyla birlikte keşfedildi, ilk kez H. habilis iskelet anatomisinin özelliklerini kesin olarak belirledi ve Homo benzerinden daha fazla Australopithecus benzeri özellikleri olduğunu ortaya çıkardı.[8] Bu nedenle, diş adaptasyonlarındaki benzerliklerin yanı sıra, Wood ve biyolojik antropolog Mark Collard, türün 1999'da Australopithecus'a taşınmasını önerdi.[12][13][14][15] Bununla birlikte, OH 62'nin daha insansı bir fizyolojiyle yeniden değerlendirilmesi, eğer doğruysa, bu konuda şüphe uyandırır.[16] 2000'lerin başlarında, erken Homo ile bazı benzerlikler sergileyen 1.8 milyon yıllık Gürcü Dmanisi kafataslarının keşfi, H. habilis ve H. rudolfensis de dahil olmak üzere Afrika'daki tüm çağdaş erken Homo gruplarının, H. erectus'a atanması gerekebileceğini gösterdi.[17][18]
Sınıflandırma
| |||||||||||||||||||||||||||||||||||||||||||||
| İnsan soyunun kök üyeleri olarak H. habilis ve H. rudolfensis'i gösteren, Homo aile ağacı.[19] |
H. habilis'in H. ergaster / H. erectus'un atası olup olmadığı veya insan (Homo) soyunun bir dalı olup olmadığı ve H. habilis'e atanan tüm örneklerin – farklı Australopithecus ve Homo türlerinin bir topluluğuna ait olabilir – doğru bir şekilde atanıp atanmadığı konusunda hala geniş bir fikir birliği yoktur.[20][21] Bununla birlikte, H. habilis ve H. rudolfensis genellikle, eşanlamlılaştırma veya cinsten çıkarma argümanları yaygın olarak benimsenmeyen, aile ağacının tabanındaki, cinsin tanınmış üyeleridir.[22]
Homo'nun Australopithecus'tan evrimleştiği konusunda büyük ölçüde fikir birliğine varılsa da, bu bölünmenin zamanlaması ve yerleşimi çok tartışıldı ve birçok Australopithecus türü ata olarak öne sürüldü. Etiyopya'nın Afar Bölgesi'nde 2,8 myö'ye tarihlenen en eski Homo örneği olan LD 350-1'in keşfi, cinsin bu zamanlarda A. afarensis'ten evrimleştiğini gösterebilir. LD 350-1'in ait olduğu tür, H. habilis'in atası olabilir, ancak bu belirsizdir.[23] En eski H. habilis örneği, AL 666-1, 2.3 myö'ye tarihlenir, ancak anatomik olarak daha genç OH 7'den daha fazla türetilmiştir (daha az bazal özelliklere sahiptir). Türetilmiş ve bazal formların aynı anda yaşadığı dönemde, H. habilis soyu 2.3 milyon yıl önce başladı.[24] Çin'in Shangchen kentinden gelen 2,1 milyon yıllık taş aletlere dayanarak, H. habilis veya bir ata türü Asya'ya dağılmış olabilir.[25] En genç H. habilis örneği, OH 13, yaklaşık 1.65 myö'ye tarihlenmektedir.[24]
Anatomi
Kafatası
Homo habilis'in beyin boyutunun Homo erectus ve Homo ergaster'dan daha küçük olmasıyla birlikte, beyin boyutunun insan evrimi çizgisi boyunca türler arasındaki geçişte özellikle hızlı bir şekilde arttığı düşünülmüştür. Homo habilis'te yaklaşık 600-650 cc'den beyin büyüklüğü, H. ergaster ve H. erectus'ta yaklaşık 900-1.000 cc'ye sıçramaktadır.[24][26] Bununla birlikte, 2015 yılında yapılan bir araştırma, OH 7'nin beyin hacmi 647-687 cc'den 729-824 cc'ye yeniden değerlendirildikten sonra H. habilis, H. rudolfensis ve H. ergaster'in beyin boyutlarının genellikle 500-900 cc arasında değiştiğini göstermiştir.[24] Yine de bu, genellikle 400-500 cc arasında değişen australopithecine beyin büyüklüğünden bir sıçramayı gösterir.[26]
Tüm Homo türlerinin beyin anatomisi, australopitesinlere kıyasla genişlemiş bir uç beyine sahiptir. OH 65'in sağa eğik dişlerinde, kişinin bir et parçasını sağ elini kullanarak bir taş aletle kesmeye çalışırken dişleri ve sol eliyle çekmesi sırasında yanlışlıkla kendi kendine oluşmuş olabilecek çizgi deseni bulunmaktadır. Doğruysa, bu sağlaklığı gösterebilir ve ellilik, beynin büyük yeniden düzenlenmesi ve sol ve sağ beyin yarımküreleri arasındaki beyin fonksiyonunun yanallaşması ile ilişkilidir. Bu senaryo, bazı Neandertal örnekleri için de varsayılmıştır. Yanallaşma, alet kullanımıyla ilişkilendirilebilir. Modern insanlarda yanallaşma dil ile zayıf bir şekilde ilişkilidir.[27]
H. habilis'in diş sıraları, daha sonraki Homo'da bulunan U-şeklinin aksine V-şeklindeydi ve burundan yukarısı düz olmasına rağmen ağız dışa doğru çıkıntılıydı (prognatikti).[24]
Yapı
OH 62 (dişi olduğu varsayılan) ve KNM-ER 3735 (erkek olduğu varsayılan) parçalı iskeletlere dayanarak, H. habilis vücut anatomisinin genellikle daha önceki A. afarensis'inkinden bile daha maymunsu olduğu ve en azından australopithecine'lerde varsayıldığı gibi ağaçlarda kısmen arboreal yaşam tarzı. OH 62'ye dayanarak ve australopithecinler ile karşılaştırılabilir vücut boyutları varsayılarak, H. habilis genellikle australopitesinler gibi küçük boyutlu olarak yorumlanmıştır, bu görüşe bağlı olarak OH 62 örneği yaklaşık 100-120 cm (3 ft 3 in-3 ft 11 in) yüksekliğinde ve 20-37 kg (44-82 lb) ağırlığında olduğu hesaplandı. Bununla birlikte, daha uzun, modern insan benzeri bacakları olduğunu varsayarsak, OH 62 yaklaşık 148 cm (4 ft 10 in) boyunda ve 35 kg (77 lb) ağırlığındadır, KNM-ER 3735 de yaklaşık aynı boyuttadır.[28] Karşılaştırma için, 1900 yılında modern insan erkek ve kadınları sırasıyla ortalama 163 cm ve 152.7 cm idi.[29] H. habilis de dahil olmak üzere H. ergaster öncesi homininlerin, genellikle erkeklerin dişilerden belirgin şekilde daha büyük olduğu ve dikkate değer bir eşeysel dimorfizm sergilediği varsayılır. Bununla birlikte, bu türde göreceli kadın vücut kütlesi bilinmemektedir.[30]
H. habilis de dahil olmak üzere erken homininlerin, modern insan dışındaki insansı maymunlar gibi kalın vücut kıllarına sahip oldukları düşünülür çünkü daha serin bölgelerde yaşıyor gibi görünüyorlar, ayrıca H. ergaster sonrası türlere göre (tüysüz olduğu varsayılan) daha az etkin bir yaşam tarzına sahip oldukları düşünülmektedir. Sonuç olarak, muhtemelen sıcak kalmak için kalın vücut kıllarına ihtiyaç duyuyorlardı.[31] Diş gelişimi oranlarına dayanarak, H. habilis'in modern insanlara kıyasla, daha çok modern insan olmayan insansı maymunlarınkine benzer şekilde, hızlandırılmış bir büyüme oranına sahip olduğu varsayılmaktadır.[32]
Uzuvlar
H. habilis ve australopitesinlerin kollarının genellikle orantılı olarak uzun olduğu ve bu nedenle tırmanma ve sallanma için uyarlanmış olduğu düşünülmüştür.[33][34][35] 2004'te antropologlar Martin Haeusler ve Henry McHenry, OH 62'nin humerus/femur oranı modern insanlar için varyasyon aralığında olduğu ve KNM-ER 3735'in modern insan ortalamasına yakın olduğu için, insansı maymun benzeri oranları olduğunu varsaymanın güvendirici olmadığını savundular. Bununla birlikte, OH 62'nin humerusu 258–270 mm (10,2–10,6 inç) uzunluğunda ve ulnası (ön kol) 245–255 mm (9,6–10,0 inç) olarak ölçülmüştür ve bu oran şempanzelerde görülen orana daha yakındır. OH 7'nin el kemikleri, el becerisi açısından önemli olan hassas kavramanın yanı sıra tırmanma için uyarlamalar önerir. Femurla ilgili olarak, geleneksel olarak A. afarensis numunesi AL 288-1 ile karşılaştırmalar, H. habilis için kalın bacakları yeniden yapılandırmak için kullanılmıştır, ancak Haeusler ve McHenry, daha ince OH 24 femurunun (H. erectus / H. ergaster veya P. boisei'ye ait) daha uygun bir karşılaştırma olabileceğini öne sürmüştür. Bu durumda, H. habilis'in daha uzun, insansı bacaklara sahip olması ve H. ergaster'de olduğu gibi, uzun mesafelerde etkili yolcular olması gerekirdi.[16] Ancak, bir fosilin korunmamış uzunluğunu tahmin etmek oldukça problemlidir. OH 62'deki uzuv kemiklerinin kalınlığı, H. ergaster / H. erectus ve modern insanlardan daha çok şempanzelere benzer.[36] OH 35'in güçlü fibulası (bu P. boisei'ye ait olsa da) daha çok insan olmayan maymunlarınkine benzer ve ağaçsıllık ve dikey tırmanma ile tutarlıdır.[37]

Bir ayak olan OH 8, karasal hareket için A. afarensis'in ayağından daha uygundur, ancak yine de tırmanma ile uyumlu birçok insansı maymun benzeri özelliği muhafaza eder.[16] Bununla birlikte, ayak, ön ayakta olduğu kadar ayak ve ayak bileği arasındaki dönüşü kısıtlayan ayak parmağı kemiği ve sıkıştırılmış orta ayak eklem yapılarına sahiptir. Ayak stabilitesi, bacak ve ayak arasındaki kuvvet transferinin etkinliğini arttırır ve bunun tersi de geçerlidir ve koşarken (ama yürüme sırasında değil) enerji üreten plantar ark elastik yay mekanizmasında rol oynar. Bu, muhtemelen H. habilis'in bir dereceye kadar dayanıklılık koşusu yapabildiğini gösterebilir; bunun tipik olarak daha sonra H. ergaster / H. erectus'ta evrimleştiği düşünülür.[38]
Diyet
H. habilis'in eti avlanmaktan ziyade çöpçülükten elde ettiği (çöpçü hipotezi), çatışmacı bir çöpçü olarak hareket ettiği ve çakallar veya çitalar gibi daha küçük yırtıcılardan avladığı düşünülmektedir.[39] Meyve muhtemelen ayrıca asitliğe tekrar tekrar maruz kalma ile tutarlı diş erozyonu ile gösterilen önemli bir diyet bileşeniydi.[40] Dental mikro aşınma-doku analizine dayanarak, H. habilis (diğer erken Homolar gibi) muhtemelen düzenli olarak sert gıdalar tüketmiyordu. Mikro giyim-doku karmaşıklığı, ortalama olarak, sert gıda yiyiciler ve yaprak yiyiciler (folivorlar) arasında bir yerdedir ve giderek genelleşen ve her yerde beslenen bir diyete işaret eder.[41][42]
Tipik olarak H. habilis ve diğer erken Homo'ların diyetlerinin Australopithecus'tan daha fazla ete sahip olduğu ve bunun beyin büyümesine yol açtığı düşünülmektedir. Bununla ilgili ana hipotezler şunlardır: et, enerji ve besin açısından zengindir ve stratejik süpürmeyi kolaylaştırmak ve taze karkasları tekelleştirmek için gelişmiş bilişsel becerilerin geliştirilmesine evrimsel baskı uygular veya et, büyük ve kalorisi pahalı insansı maymun bağırsağının boyutunun küçülmesine izin vererek buna izin verir. enerji beyin büyümesine yönlendirilir. Alternatif olarak, erken Homo'nun, yiyecek seçeneklerinin kıt olduğu kuru bir iklimde, öncelikle yeraltı depolama organlarına (yumrular gibi) ve hem erkek hem de kadın grup üyeleri arasında sosyal bağları kolaylaştıran yiyecek paylaşımına dayandığı da ileri sürülmektedir. Bununla birlikte, H. ergaster ve daha sonra Homo için tahmin edilenin aksine, kısa boylu erken Homo'nun genellikle koşmaya ve avlanmaya dayanıklı olmadığı kabul edilir ve H. habilis'in uzun ve Australopithecus benzeri ön kolu, erken Homo'nun hala bir dereceye kadar ağaçlarda yaşadığını gösterebilir. Ayrıca organize avcılık ve toplayıcılığın H. ergaster'de ortaya çıktığı düşünülmektedir. Bununla birlikte, büyük beyin büyümesini açıklamak için önerilen yiyecek toplama modelleri, günlük seyahat mesafesinin artmasını gerektirir.[43] H. habilis'in bunun yerine uzun, modern insan benzeri bacaklara sahip olduğu ve en azından kısmen ağaçta kalırken tam olarak uzun mesafeli seyahat etme yeteneğine sahip olduğu iddia edilmiştir.[16]
Australopithecus atalarına kıyasla H. habilis'in büyük kesici diş boyutu, bu türün kesici dişlere daha fazla dayandığını ima eder. H. habilis ve diğer erken Homo türlerinin alt çenelerinin gövdeleri, modern insanların ve tüm yaşayan insansı maymunlarınkinden daha kalındır, Australopithecus ile daha karşılaştırılabilir. Mandibular gövde, ısırma kuvveti veya çiğneme nedeniyle burulmaya karşı direnç gösterir, bu da çenelerinin yemek yerken alışılmadık derecede güçlü stresler üretebileceği anlamına gelir. Australopithecus'a kıyasla H. habilis'teki daha büyük molar uç kabartması, eski aletlerin sert yiyecekleri (esnek bitki parçaları veya et gibi) kırmak için kullanıldığını, aksi takdirde dişlerin daha fazla aşınacağını düşündürür. Bununla birlikte, mekanik olarak zorlayıcı gıdaları işlemek için çene uyarlamaları, teknolojik ilerlemenin diyeti büyük ölçüde etkilemediğini göstermektedir.[30]
Teknoloji

H. habilis, Erken Taş Devri Oldowan taş alet endüstrisi ile ilişkilidir. Bireyler muhtemelen bu aletleri öncelikle hayvanları kesmek, derilerini kesmek ve kemikleri ezmek için değil, aynı zamanda bazen odunları kesip kazımak ve yumuşak bitkileri kesmek için de kullandılar. Knappers'ın özenle seçilmiş litik çekirdeklere sahip olduğu ve belirli kayaların yeterince sert ve doğru yerde vurulduğunda belirli bir şekilde kırılacağını bildikleri ve kıyıcılar, polihedronlar ve diskoidler dahil olmak üzere birkaç farklı tür ürettikleri görülüyor. Bununla birlikte, belirli şekiller muhtemelen önceden düşünülmemiştir ve muhtemelen bu tür aletlerin yanı sıra yontma ustalarının emrindeki hammadde türlerinin üretiminde standardizasyon eksikliğinden kaynaklanmaktadır.[5][44] Örneğin, sferoidler, bol miktarda büyük ve yumuşak kuvars ve kuvarsit parçaları içeren Olduvai'de yaygındır, oysa Koobi Fora sferoidlerden yoksundur ve ağırlıklı olarak sert bazalt lav kayaları sağlar. H. ergaster /H. erectus tarafından icat edilen daha sonraki Aşölyen kültüründen farklı olarak, Oldowan teknolojisi planlama ve üretim için öngörü gerektirmez ve bu nedenle Oldowan taş yontucularında yüksek biliş göstermez, ancak bir dereceye kadar koordinasyon ve mekanik bilgisi gerektirir. Oldowan aletleri nadiren rötuş sergilerler ve muhtemelen çoğu zaman kullanımdan hemen sonra atılırlar.[44]

Olduwan ilk olarak 1934'te rapor edildi, ancak 1960'lara kadar, 1.8 mya'ya dayanan ve H. habilis tarafından üretilmiş en eski kültür olarak yaygın olarak kabul edilmedi. O zamandan beri, daha fazla keşif, maddi kültürün kökenlerini zaman içinde önemli ölçüde geriye yerleştirdi; Oldowan, Etiyopya'daki Ledi-Geraru ve Gona'da keşfedildi, belki de cinsin evrimi ile ilişkili olarak 2.6 milyon yıl öncesine tarihlendi (bkz. Australopithecus garhi).[5][45][5] Australopithecine'lerin ayrıca 3.3 milyon yıl önce Lomekwi taş alet endüstrisi[46] gibi aletler ürettikleri ve yaklaşık 3.4 mya'dan bazı kasaplık kanıtlarına sahip oldukları bilinmektedir.[47] Bununla birlikte, nispeten keskin kenarlı Oldowan kültürü, australopithecine teknolojisinden gelen büyük bir yenilikti ve farklı beslenme stratejilerine ve daha geniş bir gıda yelpazesini işleme yeteneğine izin verecekti, bu da zamanın değişen ikliminde avantajlı olacaktı.[45] Oldowan'ın bağımsız olarak mı icat edildiği yoksa yüz binlerce yıl boyunca birden fazla türde kayalarla yapılan hominin deneylerinin sonucu mu olduğu belli değil.[5]
1962'de Olduvai Gorge'da volkanik kayalardan yapılmış 366 cm × 427 cm × 30 cm (12 ft × 14 ft × 1 ft) daire keşfedildi. 61–76 cm (2-2,5 ft) aralıklarla, kayalar 15–23 cm (6-9 inç) yüksekliğe kadar yığılmıştır. Mary Leakey, kaya yığınlarının, muhtemelen bir rüzgar siperini veya kaba bir kulübeyi desteklemek için yere yapışmış direkleri desteklemek için kullanıldığını öne sürdü. Günümüzün bazı göçebe kabileleri, geçici barınaklar inşa etmek için benzer alçakta uzanan kaya duvarları inşa ediyor, dik dalları direkt olarak büküyor ve çimleri veya hayvan postlarını perde olarak kullanıyor.[48] Bu, 1.75 mya'ya tarihlenen, bazı erken Homo'lara atfedilir ve mimarinin iddia edilen en eski kanıtıdır.[49]
Ayrıca bakınız
Kaynakça
- ^ Antón, S. C. (2012). "Early Homo: Who, When, and Where". Current Anthropology. 53 (6): 279. doi:10.1086/667695.
- ^ Dawkins, Richard (2008). Ataların hikayesi : yaşamın kökenine yolculuk = The ancestor's tale : a pilgrimage to the dawn of evolution. Ahmet Fethi. İstanbul: Hil Yayın. s. 87. ISBN 978-975-7638-34-6. OCLC 263055964.
- ^ a b c d "Homo habilis—A Premature Discovery: Remembered by One of Its Founding Fathers, 42 Years Later". The First Humans – Origin and Early Evolution of the Genus Homo. Vertebrate Paleobiology and Paleoanthropology. Springer, Dordrecht. 2006. ss. 7-15. doi:10.1007/978-1-4020-9980-9_2. ISBN 978-1-4020-9980-9.
- ^ a b Leakey (1964). "A New Species of the Genus Homo from Olduvai Gorge" (PDF). Nature. 202 (4927): 7-9. doi:10.1038/202007a0. PMID 14166722. 1 Mart 2021 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 12 Ekim 2021.
- ^ a b c d e de la Torre (2011). "The origins of stone tool technology in Africa: a historical perspective". Philosophical Transactions of the Royal Society B. 366 (1567): 1028-1037. doi:10.1098/rstb.2010.0350. PMC 3049100 $2. PMID 21357225.
- ^ Wood (2000). "Human evolution: taxonomy and paleobiology". Journal of Anatomy. 197: 39-41. doi:10.1046/j.1469-7580.2000.19710019.x. PMC 1468107 $2. PMID 10999270.
- ^ a b Tobias (1983). "Hominid evolution in Africa" (PDF). Canadian Journal of Anthropology. 3 (2): 163-183. 7 Temmuz 2021 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 12 Ekim 2021.
- ^ a b Johanson (1987). "New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania". Nature. 327 (6119): 205-209. doi:10.1038/327205a0. PMID 3106831.
- ^ Delson, E., (Ed.) (1985). "Early Homo in Kenya, systematic relationships". Ancestors: The hard evidence. Alan R. Liss. ISBN 978-0-8451-0249-7.
- ^ Wood (1999). "Homo rudolfensis Alexeev, 1986: Fact or phantom?". Journal of Human Evolution. 36 (1): 115-118. doi:10.1006/jhev.1998.0246. PMID 9924136.
- ^ Curnoe (2010). "A review of early Homo in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (Homo gautengensis sp. nov.)". HOMO - Journal of Comparative Human Biology. 61 (3): 151-177. doi:10.1016/j.jchb.2010.04.002. PMID 20466364. 1 Haziran 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ekim 2021.
- ^ Wood (1999). "The Human Genus" (PDF). Science. 284 (5411): 65-71. doi:10.1126/science.284.5411.65. PMID 10102822. 23 Kasım 2020 tarihinde kaynağından (PDF) arşivlendi.
- ^ Miller J. M. A. (2000). "Craniofacial variation in Homo habilis: an analysis of the evidence for multiple species". American Journal of Physical Anthropology. 112 (1): 103-128. doi:10.1002/(SICI)1096-8644(200005)112:1<103::AID-AJPA10>3.0.CO;2-6. PMID 10766947.
- ^ Tobias (1991). "The species Homo habilis: example of a premature discovery". Annales Zoologici Fennici. 28 (3–4): 371-380.
- ^ "Defining the Genus Homo". Handbook of Paleoanthropology. 2015. ss. 2107-2144. doi:10.1007/978-3-642-39979-4_51. ISBN 978-3-642-39978-7.
- ^ a b c d Haeusler (2004). "Body Proportions of Homo Habilis". Journal of Human Evolution. 46 (4): 433-465. doi:10.1016/j.jhevol.2004.01.004. PMID 15066379.
- ^ Margvelashvili (2013). "Tooth wear and dentoalveolar remodeling are key factors of morphological variation in the Dmanisi mandibles". Proceedings of the National Academy of Sciences (İngilizce). 110 (43): 17278-83. doi:10.1073/pnas.1316052110. ISSN 0027-8424. PMC 3808665 $2. PMID 24101504.
- ^ Lordkipanidze (2013). "A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo". Science (İngilizce). 342 (6156): 326-331. doi:10.1126/science.1238484. ISSN 0036-8075. PMID 24136960.
- ^ Strait, D.; Grine, F.; Fleagle, J. G. (2015). "Analyzing Hominin Phylogeny: Cladistic Approach" (PDF). Henke, W.; Tattersall, I. (Ed.). Handbook of Paleoanthropology. 2nd. Springer. s. 2006. doi:10.1007/978-3-642-39979-4_58. ISBN 978-3-642-39979-4.
- ^ Tattersall (2019). "Classification and phylogeny in human evolution". Ludus Vitalis. 9 (15): 139-140. 15 Nisan 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ekim 2021.
- ^ Extinct Humans. Basic Books. 2001. s. 111. ISBN 978-0-8133-3918-4.
- ^ "Analyzing Hominin Phylogeny: Cladistic Approach". Handbook of Paleoanthropology. 2nd. Springer. 2014. ss. 2005-2006. ISBN 978-3-642-39979-4.
- ^ Villmoare (2015). "Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia". Science. 347 (6228): 1352-1355. doi:10.1126/science.aaa1343. PMID 25739410.
- ^ a b c d e F. Spoor (2015). "Reconstructed Homo habilis type OH 7 suggests deep-rooted species diversity in early Homo". Nature. 519 (7541): 83-86. doi:10.1038/nature14224. PMID 25739632.
- ^ Zhu (2018). "Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago". Nature. 559 (7715): 608-612. doi:10.1038/s41586-018-0299-4. PMID 29995848.
- ^ a b Tobias (1987). "The brain of Homo habilis: A new level of organization in cerebral evolution". Journal of Human Evolution. 16 (7–8): 741-761. doi:10.1016/0047-2484(87)90022-4.
- ^ Frayer (2016). "OH-65: The earliest evidence for right-handedness in the fossil record". Journal of Human Evolution. 100: 65-72. doi:10.1016/j.jhevol.2016.07.002. PMID 27765150.
- ^ Will (2015). "Spatial and temporal variation of body size among early Homo". Journal of Human Evolution. 82: 15-33. doi:10.1016/j.jhevol.2015.02.009. PMID 25818180.
- ^ Roser (2013). "Human Height". Our World in Data. 30 Ocak 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 16 Haziran 2020.
- ^ a b Ungar (2006). "Diet in Early Homo: A Review of the Evidence and a New Model of Adaptive Versatility". Annual Review of Anthropology. 35: 208-228. doi:10.1146/annurev.anthro.35.081705.123153.
- ^ Dávid-Berrett (2016). "Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling". Journal of Human Evolution. 94: 72-82. doi:10.1016/j.jhevol.2016.02.006. PMC 4874949 $2. PMID 27178459.
- ^ Schwartz (2012). "Growth, Development, and Life History throughout the Evolution of Homo". Current Anthropology. 53 (6): 400-401. doi:10.1086/667591.
- ^ Haeusler (2007). "Evolutionary reversals of limb proportions in early hominids? evidence from KNM-ER 3735 (Homo habilis)". Journal of Human Evolution. 53 (4): 385-405. doi:10.1016/j.jhevol.2007.06.001. PMID 17688910.
- ^ Donald C. Johanson (21 Mayıs 1987). "New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania". Nature. 327 (6119): 205-209. doi:10.1038/327205a0. PMID 3106831.
- ^ Wood, B. (1987). "Who is the 'real' Homo habilis?". Nature. 327 (6119): 187-188. doi:10.1038/327187a0. PMID 3106828.
- ^ Ruff (2009). "Relative Limb Strength and Locomotion in Homo habilis". American Journal of Physical Anthropology. 138 (1): 90-100. doi:10.1002/ajpa.20907. PMID 18711733.
- ^ Marchi (2019). "Relative fibular strength and locomotor behavior in KNM-WT 15000 and OH 35". Journal of Human Evolution. 131: 48-60. doi:10.1016/j.jhevol.2019.02.005. PMID 31182206.
- ^ Bramble (2004). "Endurance running and the evolution of Homo". Nature. 432 (7015): 345-352. doi:10.1038/nature03052. PMID 15549097.
- ^ Cavallo (1989). "Tree-stored leopard kills: expanding the hominid scavenging niche". Journal of Human Evolution. 18 (4): 393-399. doi:10.1016/0047-2484(89)90038-9.
- ^ Peuch (1984). "Acidic-Food Choice in Homo habilis at Olduvai". Current Anthropology. 25 (3): 349-350. doi:10.1086/203146.
- ^ Ungar (9 Şubat 2012). "Dental Evidence for the Reconstruction of Diet in African Early Homo". Current Anthropology. 53: S318-S329. doi:10.1086/666700.
- ^ Ungar (1 Ocak 2006). "Dental Microwear and Diets of African Early Homo". Journal of Human Evolution. 50 (1): 78-95. doi:10.1016/j.jhevol.2005.08.007. PMID 16226788.
- ^ Pontzer (2012). "Ecological Energetics in Early Homo". Current Anthropology. 56 (6): 346-358. doi:10.1086/667402.
- ^ a b Toth (1985). "The oldowan reassessed: A close look at early stone artifacts". Journal of Archaeological Science. 12 (2): 101-120. doi:10.1016/0305-4403(85)90056-1.
- ^ a b Braun (2019). "Earliest known Oldowan artifacts at >2.58 Ma from Ledi-Geraru, Ethiopia, highlight early technological diversity". Proceedings of the National Academy of Sciences. 116 (24): 11712-11717. doi:10.1073/pnas.1820177116. PMC 6575601 $2. PMID 31160451.
- ^ Harmand (2015). "3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya". Nature. 521 (7552): 310-315. doi:10.1038/nature14464. PMID 25993961.
- ^ McPherron (2010). "Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia". Nature. 466 (7308): 857-860. doi:10.1038/nature09248. PMID 20703305.
- ^ Olduvai Gorge: Volume 3, Excavations in Beds I and II, 1960-1963. Cambridge University Press. 1971. s. 24. ISBN 9780521077231. 7 Nisan 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 21 Ağustos 2022.
- ^ "Building, dwelling, living: how animals and people make themselves at home in the world". The Perception of the Environment: Essays on Livelihood, Dwelling and Skill. Psychology Press. 2000. s. 184. ISBN 9780415228329.
Dış bağlantılar
- John Gurche tarafından H. habilis'in rekonstrüksiyonları 6 Eylül 2021 tarihinde Wayback Machine sitesinde arşivlendi.
- Arkeoloji Bilgisi 26 Mayıs 2011 tarihinde Wayback Machine sitesinde arşivlendi.
- Homo habilis 12 Ekim 2021 tarihinde Wayback Machine sitesinde arşivlendi. – Smithsonian Enstitüsü'nün İnsan Kökenleri Programı
- İnsan Zaman Çizelgesi (Etkileşimli) 17 Temmuz 2017 tarihinde Wayback Machine sitesinde arşivlendi. – Smithsonian, Ulusal Doğa Tarihi Müzesi (Ağustos 2016).