
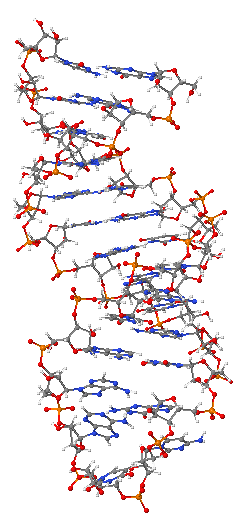
Deoksiriboz nükleik asit veya kısaca DNA, tüm organizmaların ve bazı virüslerin canlılık işlevleri ve biyolojik gelişmeleri için gerekli olan genetik talimatları taşıyan bir nükleik asittir. DNA'nın başlıca rolü bilgiyi uzun süre saklamasıdır. Protein ve RNA gibi hücrenin diğer bileşenlerinin inşası için gerekli olan bilgileri içermesinden dolayı DNA; bir kalıp, şablon veya reçeteye benzetilir. Bu genetik bilgileri içeren DNA parçaları gen olarak adlandırılır. Bazı DNA dizilerinin yapısal işlevleri vardır, diğerleri ise bu genetik bilginin ne şekilde kullanılacağının düzenlenmesine yararlar.

Bakteri (İngilizce telaffuz: [bækˈtɪəriə] ( dinle); tekil isim: bacterium), tek hücreli mikroorganizma grubudur. Tipik olarak birkaç mikrometre uzunluğunda olan bakterilerin çeşitli şekilleri vardır, kimi küresel, kimi spiral şekilli, kimi çubuksu, kimi virgül şeklinde olabilir. Yeryüzündeki her ortamda bakteriler mevcuttur. Toprakta, deniz suyunda, okyanusun derinliklerinde, yer kabuğunda, deride, hayvanların bağırsaklarında, asitli sıcak su kaynaklarında, radyoaktif atıklarda büyüyebilen tipleri vardır. Tipik olarak bir gram toprakta bulunan bakteri hücrelerinin sayısı 40 milyon, bir mililitre tatlı suda ise bir milyondur; toplu olarak dünyada beş nonilyon (5×1030) bakteri bulunmaktadır, bunlar dünyadaki biyokütlenin çoğunu oluşturur. Bakteriler gıdaların geri dönüşümü için hayati bir öneme sahiptirler ve gıda döngülerindeki çoğu önemli adım, atmosferden azot fiksasyonu gibi, bakterilere bağlıdır. Ancak bu bakterilerin çoğu henüz tanımlanmamıştır ve bakteri şubelerinin sadece yaklaşık yarısı laboratuvarda kültürlenebilen türlere sahiptir. Bakterilerin araştırıldığı bilim bakteriyolojidir, bu, mikrobiyolojinin bir dalıdır.
dinle); tekil isim: bacterium), tek hücreli mikroorganizma grubudur. Tipik olarak birkaç mikrometre uzunluğunda olan bakterilerin çeşitli şekilleri vardır, kimi küresel, kimi spiral şekilli, kimi çubuksu, kimi virgül şeklinde olabilir. Yeryüzündeki her ortamda bakteriler mevcuttur. Toprakta, deniz suyunda, okyanusun derinliklerinde, yer kabuğunda, deride, hayvanların bağırsaklarında, asitli sıcak su kaynaklarında, radyoaktif atıklarda büyüyebilen tipleri vardır. Tipik olarak bir gram toprakta bulunan bakteri hücrelerinin sayısı 40 milyon, bir mililitre tatlı suda ise bir milyondur; toplu olarak dünyada beş nonilyon (5×1030) bakteri bulunmaktadır, bunlar dünyadaki biyokütlenin çoğunu oluşturur. Bakteriler gıdaların geri dönüşümü için hayati bir öneme sahiptirler ve gıda döngülerindeki çoğu önemli adım, atmosferden azot fiksasyonu gibi, bakterilere bağlıdır. Ancak bu bakterilerin çoğu henüz tanımlanmamıştır ve bakteri şubelerinin sadece yaklaşık yarısı laboratuvarda kültürlenebilen türlere sahiptir. Bakterilerin araştırıldığı bilim bakteriyolojidir, bu, mikrobiyolojinin bir dalıdır.
Ribonükleik asid (RNA), bir nükleik asittir, nükleotitlerden oluşan bir polimerdir. Her nükleotit bir azotlu baz, bir riboz şeker ve bir fosfattan oluşur. RNA pek çok önemli biyolojik rol oynar, DNA'da taşınan genetik bilginin proteine çevirisi (translasyon) ile ilişkili çeşitli süreçlerde de yer alır. RNA tiplerinden olan mesajcı RNA, DNA'daki bilgiyi protein sentez yeri olan ribozomlara taşır, ribozomal RNA ribozomun en önemli kısımlarını oluşturur, taşıyıcı RNA ise protein sentezinde kullanılmak üzere kullanılacak aminoasitlerin taşınmasında gereklidir. Ayrıca çeşitli RNA tipleri genlerin ne derece aktif olduğunu düzenlemeye yarar.

Crosover veya krossing over ya da parça değişimi mayoz bölünmenin profaz I evresinde görülen, çift halde bulunan kromozomların yaptığı parça değişimine verilen addır. Bunun sonucunda genetik rekombinasyon meydana gelir. Yani farklı kromozomlarda bulunan genlerin alelleri birbiriyle yer değiştirir.

Genetikte, mikroRNA (miRNA) yaklaşık 21-23 nükleotit uzunluğunda tek iplikli RNA molekülü türüdür, gen ifadesinin düzenlenmesinde rol oynar. miRNA'lar kodlamayan RNA'lardandır, yani DNA'dan transkripsiyonu yapılan ama proteine çevirisi yapılmayan genler tarafından kodlanırlar. Pri-miRNA olarak adlandırılan primer transkriptler işlenerek, önce pre-miRNA adlı kısa sap-ilmik yapılarına, sonra da fonksiyonel miRNA'ya dönüşürler. Olgun miRNA moleküller bir veya daha çok mesajcı RNA (mRNA) ile kısmî tamamlayıcıdır ve başlıca işlevleri gen ifadesini aşağı ayarlamaktır. 1993'te Lee ve çalışma arkadaşları tarafından Victor Ambros laboratuvarında keşfedilmişlerdir, ancak mikroRNA terimi ilk 2001'de kullanıma girimiştir.

DNA polimeraz, DNA replikasyonunu sağlayan bir enzimdir. Bu enzimler bir DNA ipliğini kalıp olarak kullanır, onu okuyup, onun boyunca deoksiribonükleotitlerin polimerizasyonunu katalizler. Yeni polimerleşmiş molekül kalıp ipliği tamamlayıcıdır ve kalıp ipliğin eski eşi ile aynı yapıya sahiptir.
Genetik çeşitlenme veya rekombinasyon, genetik materyalin bir zincirinin kırılması ve sonrasında farklı bir DNA molekülüne katılmasıyla oluşan süreçtir. Ökaryotlarda çeşitlilik genellikle mayoz sırasında, kromozom çiftleri arasındaki kromozomal parça değişimiyle meydana gelir. Bu süreç döllerin ebeveynlerinden farklı gen dizilimlerine sahip olmasına ve daha önce olmayan yeni alleler üretmesine öncülük eder.
Nükleaz, nükleik asitleri kısmen veya tamamen parçalayan bir enzim tipidir. Bu enzimler gerek sindirim sisteminde, gerek de hücre içinde, örneğin hata tamiri, gen regülasyonu, viral savunma gibi önemli işlevlerin gerçekleşmesinde rol oynarlar. Nükleazlar, tiplerine bağlı olarak, DNA ve RNA zincirlerini çeşitli biçimlerde kesebilirler. Gen mühendisliğinde farklı nükleazlar DNA moleküllerinin arzu edilen biçime sokulmasında, ayrıca DNA ve RNA moleküllerinin yapılarının anlaşılmasında birer araç olarak kullanılır.

Helikazlar tüm canlılar için hayatî önem taşıyan bir enzim sınıfıdır. Nükleik asitlerin fosfodiester omurgası üzerinde hareket ederek birbirlerine hidrojen bağlarıyla bağlanmış nükleik asit ipliklerini ayrıştırır. Bunun için ATP hidrolizinden açığa çıkan enerjiyi kullanır.
DNA yapısı, hem tek iplikli hem çift iplikli DNA'da çeşitli biçimler gösterir. Hücreler için DNA'nın yapısıyla ilişkili olan DNA'nın mekanik yapısı hücreler için önemli bir sorun yaratır. DNA'nın okunması veya ona bağlanmasıyla ilgili her hücresel süreç, onun tanınması, paketlenmesi veya değişime uğratılmasına etki edecek şekilde onun mekanik yapılarını da kullanır ya da değiştirir. DNA 'nın aşırı uzunluğunun, onun sertliğinin ve sarmal yapısının bir sonucu olarak, hücre DNA'sının düzenlenebilmesi için histon gibi yapısal proteinler ve topoizomeraz ve helikaz gibi enzimler evrimleşmiştir. DNA'nın özellikleri onun moleküler yapısı ve dizisi ile yakından ilişkilidir. Özellikle DNA ipliklerini birbirine bağlayan hidrojen bağları ve elektronik etkileşimlerin, her bir iplikteki bağların kuvvetine kıyasla olan zayıflığı, bu ilişkide önemli bir rol oynar.

Üç iplikli DNA veya üç sarmallı DNA, üç oligonükleotitin birbiri etrafına sarılarak üçlü sarmal oluşturduğu bir DNA yapısıdır. Bu yapıda ipliklerden biri, B-biçimli DNA ile Hoogsteen veya ters Hoogsteen hidrojen bağları kurar. Bu bağ, ikili sarmalın büyük oluğunda oluşur.

Homolog rekombinasyon, benzer veya aynı dizilere sahip DNA iplikleri arasında nükleotit dizilerinin birbiriyle yer değiştirdiği bir genetik rekombinasyon tipidir. Bu süreç sırasında DNA birkaç kere kesilir, sonra da birleştirilir. Homolog rekombinasyon, DNA'daki çift iplikli kırıkların hatasız tamirinde kullanılmanın yanı sıra, mayoz sırasında krosover yoluyla yeni DNA dizi bileşimlerinin (kombinezonlarının) oluşumunu da sağlar. DNA'daki yeni bileşimler genetik varyasyonlar oluşturur. Genetik varyasyonlar yeni, bir olasılıkla yararlı olabilecek alel kombinasyonlarıdır, bunların üreyen canlı topluluklarda oluşmaları, bu değişiklikleri taşıyan bireylerin değişen çevresel şartlara evrimsel adaptasyon göstermelerini sağlar.

RAD51, bir insan genidir, bu genin ürünü olan ve aynı isme sahip olan protein DNA çift iplikçikli kesiklerini tamir eder. Bu protein RAD51 protein ailesinin üyelerinden biridir. RAD51 protein ailesi üyeleri bakteriyel RecA ve maya Rad51 ile homologdur. Protein, mayadan insana kadar çoğu ökaryotta korunmuştur.
Mikrosatelitler, Basit dizi tekrarları veya Kısa Bitişik Tekrarlar DNA'da bulunan, 1-6 baz çifti uzunluğundaki tekrar eden dizilerdir.

T7 RNA Polymeraz T7 bakteriyofaja ait bir RNA polimerazdır. Bu enzim, faj genlerindeki genetik bilginin mesajcı RNA molekülü şeklinde transkripsiyonunu katalizler.
Chargaff kuralları, Avusturyalı biyokimyacı Edwin Chargaff tarafından 1949-1951'de yayımlanan, DNA'daki çeşitli azotlu bazların miktarları arasındaki ilişkileri ifade eden empirik kurallardır.

Phillip Allen Sharp, Amerikan genetikçi ve moleküler biyolog. RNA bağlanmasının kaşiflerinden biridir. Richard J. Roberts ile birlikte ökaryot hücrelerinin DNA dizelerindeki genlerin bitişik sırada olmadığını, aralarda intron denilen okunmayan ve protein sentezine katılmayan bölümlerin olduğunu keşfettiler. Bu sayede mRNA'lar aynı DNA dizesinden bu bölümleri farklı şekilde silmeleri ile farklı proteinleri kodlayabilmektedir. İkili bu keşifleri ile 1993 Nobel Fizyoloji veya Tıp Ödülünü kazanmışlardır.
Moleküler evrim, nesiller boyu aktarılacak şekilde, DNA, RNA ve protein gibi hücresel moleküllerin diziliminin değiştirilmesi işlemidir ya da bununla ilgilenen bilim dalıdır. Moleküler evrimin alanı, bu değişimlerdeki kalıpları açıklamak için evrimsel biyoloji ve popülasyon genetiği ilkelerini kullanır. Moleküler evrim başlıca, nükleotid değişimlerinin oranları ve etkilerini, nötr evrimi, doğal seçilimi, yeni genlerin kökenlerini, karmaşık özelliklerin genetik yapısını, türleşmenin genetik temelini, gelişim evrimini ve evrimin genomik ve fenotipik değişikliklere neden olan etkilerini inceler.
Homoloji yönlendirmeli tamir hücrede çift iplik hasarlarını tamir amacı güden mekanizma. En yaygın formu homolog rekombinasyondur. HDR, genellikle G2 ve S fazında, sadece çekirdekte tamiri yapılacak DNA' ya homolog bir DNA parçası varlığında çalışır. Eğer bu homolog parça yok ise homolog olmayan uç birleştirme mekanizması devreye girer.

Homolog olmayan uç birleştirme çift zincir kırığı düzeltme yolaklarından biri. NHEJ homoloji yönlendirmeli tamire karşın çift zincir kırığını herhangi ekleme yapmadan direkt olarak birleştirir. Yani herhangi bir -tamire rehberlik edecek- homolog kalıba gerek duymaz. "Homolog olmayan uç birleştirme" terimi 1996'da Moore ve Haber tarafından ortaya atılmıştır.

