Gorgonopsia
| Gorgonopsia | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
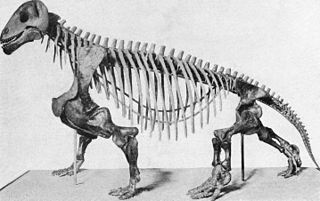
 Lycaenops ornatus'un montelenmiş iskeleti, Amerikan Doğa Tarihi Müzesi | |||||||||||
| Biyolojik sınıflandırma | |||||||||||
| |||||||||||
| Tip cins | |||||||||||
| †Gorgonops Owen, 1876 | |||||||||||
| Alt gruplar | |||||||||||
Gorgonopsiyenler (Gorgonopsia),[a] Geç Permiyen'in baskın etçil terapsit grubu. Permiyen'de, kesit olarak oval olan ve tırtıklı bir arka kenar taşıyan büyük ölçüde genişlemiş üst ve alt köpek dişleri tarafından av üzerinde beslenmeye uyum sağlamış, nispeten nadir, küçük etoburlar olarak ortaya çıkarlar. Bununla birlikte, Geç Permiyen'in geri kalanında, gorgonopsiyenler baskın karasal etoburlardı ve aynı zamanda çağdaş Rus yataklarında da bulunurlar. Şimdiye kadar Güney Afrika ve Rusya dışında tamamen bilinmiyorlar ve diğer son Permiyen terapsit gruplarının aksine, tek bir gorgonopsiyenin bile Triyas'ın sonuna kadar hayatta kaldığı bilinmiyor.[3]
Daha önce yaşamış olası terapsit olarak bahsedilen Kuzey Amerika'nın Guadalupiyeni'nden büyük ölçüde gözden düşmüş parçalanmış örnekler arasında, bir bazal gorgonopsiyen olabilecek Watongia vardır. Watongia, iki kranial fragman ve birkaç postkraniyal kemik ile temsil edilir. Olson'a göre, kafatası çatısının parçası üzerinde bir preparietal kemik ve aynı zamanda biraz gorgonopsiyen benzeri omurlara sahipti. Ayrıca ince uzuv kemikleri ve arka ayağın belirli falanjlarının boyutunda bir miktar küçülme vardı.[4]
Morfoloji

Gorgonopsiyenlerin vücut boyutları küçük bir köpeğinkinden, yaşayan herhangi bir memeli yırtıcı hayvandan biraz daha büyük olana kadar değişiyordu. Büyütülmüş köpek dişlerinin yanı sıra, beş üst ve dört alt iyi gelişmiş kesici diş bulunurken, postkaninler en fazla dört veya beş çok küçük, basit dişe indirgenir. Aşağıdakiler de dahil olmak üzere birkaç kranial özellik de grubu teşhis eder:[3]
- kafatasının genişlemiş, düz tepeli preorbital bölgesi
- intertemporal çatı geniş ve adduktor kas tarafından işgal edilmemiş halde kalırken, temporal fenestranın lateral ve posterior uzantı ile genişlemesi
- preparietal kemik
- vomerler kaynaşmış haldedir
- damağın orta ventral hattında buluşan çift palatin kemikleri, pterygoidlerle birlikte derin tonozlu bir damak oluşturur.
Sistematik
Gorgonopsiyenler ayrıca etçil dişler, kafatası çatısının geniş intertemporal bölgesi ve çok muhafazakar postkraniyal iskelet gibi birçok ilkel therapsid özelliklerini de korurlar. Sonuç olarak, geçmişte onları, şu anda Biarmosuchia'ya dahil olan hipposauridler ve burnetiidler gibi diğer ilkel etçil taksonlarla sınıflandırmaya yönelik bir eğilim olmuştur.[3]
Daha sonraki yazarlar Sigogneau-Russell'in (1989) incelemesini takip etti ve Gorgonopsia'yı, kafatasının temporal fenestrasının ve damak notunun tanısal karakterlerine sahip iyi tanımlanmış formlarla sınırlandırdı. Taksonomik olarak bu şekilde kısıtlanmış olan grup içinde dikkate değer bir muhafazakarlık vardır. Cinsler kendi aralarında büyüklük, kafatası genişliğinin nispi oranları, interorbital genişlik, vb. ve postcanine diş sayısının önemsiz karakterinden biraz daha fazla farklılık gösterir ve hepsi tek bir Gorgonopsidae familyası içinde yer alabilir.[3]
Sigogneau-Russell (1989) üç alt aileyi tanır. Gorgonopsinae cinslerin çoğunluğundan oluşur. Rubidgeinae, geniş postorbital bar ve büyük elmacık kemerleri ile çok geniş, ağır yapılı bir kafatasına sahiptir. Inostranceviinae, esas olarak, tüm gorgonopsianların en büyüğü olan ve yarım metreden fazla kafatası uzunluğuna sahip olan Rus cinsi Inostrancevia'nın kendisinden oluşur. Bu boyuttan ayrı olarak ana özelliği, nispeten çok uzun preorbital uzunluktur.[3]
Dış sistematik
Rubidge & Sidor (2001) tarafından oluşturulmuş, Gorgonopsiyenlerin dış sistematiğini gösteren kladogram:[5]
| Therapsida |
| ||||||||||||||||||||||||

İç sistematik
Bendel vd. (2018) tarafından yapılan çalışmaya göre oluşturulmuş, gorgonopsia'nın iç sistematiğini gösteren kladogram.[6]
| Gorgonopsia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiyoloji

Biyolojik olarak, gorgonopsiyenler son derece yırtıcı bir yaşam tarzına mükemmel bir şekilde adapte olmuşlardı.[3]
Çene hareketi

Gorgonopsiyenler, kafatasına sadece gevşek bir şekilde bağlı olan bir kuadrata sahiptiler (kuadrat ve skuamosal arasında sütür bağlantısı yoktu) ve çene tamamen açıldığında, çene ekleminin şekli eklemi kuadrata göre çevirmeye zorladı. (eklem çeneyi açmak için döndüğü için yana doğru da kaymaya zorlandı).[5]
Kemp (1969), çenelerin işleyişini ayrıntılı olarak analiz etti ve Leontocephalus'un olağanüstü iyi korunmuş bir örneğinin karşı kesici ve köpek dişlerinde korunan aşınma fasetlerinden, alt çenenin iki hareket tarzına sahip olduğunu gösterdi. En basiti, büyük üst ve alt köpek dişlerinin birbirini yeterince temizlemesi için yeterli olacak şekilde, çeneleri 90 derece kadar son derece geniş bir şekilde açmaktan ibaretti. Alt çene daha sonra güçlü bir şekilde eklendi, böylece keskin, tırtıklı kenarlı köpekler, onu etkisiz hale getirmek için avın içine batacaktı. İkinci ısırma şekli daha kesindi ve alt çenenin ileriye doğru bir propalinal kaymasından oluşuyordu, böylece kapatıldığında bir taraftaki dört tırtıklı alt kesici diş benzer şekilde tırtıklı beş üst kesici dişin arasından geçiyordu. İç içe geçen kesici dişin etkisi, ette pürüzlü bir kenar kesmek ve bu nedenle parçaların daha kolay yırtılıp yutulmasını sağlamak olurdu. Çenenin gerekli ön-arka hareketini elde etmek için, kuadrat ve skuamosal kemik arasında hareketli bir artikülasyon gelişti, bu sayede kuadrat kemiğinin top şeklindeki gövdesi, enine bir eksen etrafında yuva şeklinde bir skuamozal girinti içinde döndürüldü. Etki, kuadratın alt kısmının ileri veya geri kaymasına izin vermekti, bu da tüm alt çenenin ileri veya geri hareket etmesine neden oldu. Bu sayede, kesici dişlerin birbirine geçmesi için çene öne doğru veya kesici dişlerin güçlü köpek hareketinin önüne geçmemesi için geriye doğru kayabilir.[3]
Laurin (1998), gorgonopsiyenlerin kuadratının bu propalinal yeteneğe sahip olup olmadığından şüpheliydi, ancak alt çene, alt kesici dişler için yeterince uzun olmadığından, aksi takdirde kesici dişlerin yanlarındaki aşınma fasetlerinin nasıl oluşabileceği konusunda yorum yapmadı.[3]
Her halükarda, gorgonopsiyen diş yapısı, muhtemelen bol miktarda daha küçük dicynodontlar'dan oluşan nispeten büyük, aktif avlarla başa çıkmak için uyarlanmıştır.[3]
Taksonomi
- Klad † Gorgonopsia
- Familya † Gorgonopsidae
- Aelurognathus
- Aelurosaurus
- Aloposaurus
- Arctognathus
- Arctops
- Clelandina
- Cynariops
- Cyonosaurus
- Dinogorgon
- Eriphostoma
- Gorgonops
- Inostrancevia
- Leogorgon
- Leontosaurus
- Lycaenops
- Njalila
- Nochnitsa
- Pravoslavlevia
- Rubidgea
- Ruhuhucerberus
- Sauroctonus
- Smilesaurus
- Suchogorgon
- Sycosaurus
- Viatkogorgon
- Familya † Gorgonopsidae
Fosil dağılımı

Gorgonopsiyenlerin fosilleri, Güney Afrika, Zambiya, Tanzanya, Malawi ve Rusya Federasyonu'nda bulundu.[1]
Ayrıca bakınız
Notlar
- ^ Ad, Yunan mitolojisindeki Gorgonlar'dan esinlenerek türetilmiştir – ὄψις (ópsis) Antik Yunancada yüz veya görünüş anlamına gelir.[2]
Kaynakça
Dipnotlar
- ^ a b "Gorgonopsia". paleobiodb.org. 7 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Ağustos 2021.
- ^ "ὄψις İngilizce içinde - Grekçe-İngilizce Sözlük | Glosbe". glosbe.com. 9 Nisan 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Nisan 2022.
- ^ a b c d e f g h i T.S. Kemp The origin and evolution of mammals (2005) ss. 52-55.
- ^ T.S. Kemp The origin and evolution of mammals (2005) ss. 28-30.
- ^ a b Rubidge, Bruce S.; Sidor, Christian A. (Kasım 2001). "Evolutionary Patterns Among Permo-Triassic Therapsids". Annual Review of Ecology and Systematics. 32 (1): 449-480. doi:10.1146/annurev.ecolsys.32.081501.114113. ISSN 0066-4162.
- ^ Kammerer, Christian F.; Masyutin, Vladimir (8 Haziran 2018). "Gorgonopsian therapsids (Nochnitsa gen. nov. and Viatkogorgon) from the Permian Kotelnich locality of Russia". PeerJ (İngilizce). 6: e4954. doi:10.7717/peerj.4954. ISSN 2167-8359. PMC 5995105 $2. PMID 29900078. 1 Temmuz 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Ağustos 2021.
Bibliyografya
- Kemp, T. S. (2005). The origin and evolution of mammals. Oxford: Oxford University Press. ISBN 978-0-19-154517-7. OCLC 232311794. 9 Eylül 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Ağustos 2021.
Dış bağlantılar
 Wikimedia Commons'ta Gorgonopsia ile ilgili çoklu ortam belgeleri bulunur
Wikimedia Commons'ta Gorgonopsia ile ilgili çoklu ortam belgeleri bulunur Vikitür'de Gorgonopsia ile ilgili ayrıntılı taksonomik bilgiler bulunur.
Vikitür'de Gorgonopsia ile ilgili ayrıntılı taksonomik bilgiler bulunur.- Prehistoric: Therapsid & Gorgonopsia 7 Ağustos 2021 tarihinde Wayback Machine sitesinde arşivlendi. inaturalist.org
- Video: When Gorgons Rules the Earth (4:00)