Golgi aygıtı


Golgi aygıtı (ayrıca Golgi cisimciği veya Golgi kompleksi), çoğu ökaryotik hücrede bulunan bir organeldir.[1] 1897'de, Golgi aygıtı isminin kaynağı olan, İtalyan tabip Camillo Golgi tarafından keşfedilmiştir.[2]
Proteinler sentezlenip hedef noktalarına gitmeden önce golgi tarafından işlenir ve paketlenir; bu özellikle sekresyon için işlenen proteinlerde önemidir. Golgi aygıtı hücresel endomembran sisteminin bir bölümünü oluşturur.
Keşfi
Oldukça büyük boyutundan dolayı, golgi aygıtı keşfedilip ayrıntılarıyla incelenen ilk organellerden biridir. Bu aygıt ilk defa 1897 yılında İtalyan tabip Camillo Golgi tarafından sinir sistemi üzerinde yapılan bir inceleme sonucu keşfedilmiştir.[2] Bu organeli kendi mikroskobunda ilk keşfinde yapıyı apparato reticolare interno (Türkçe: dahili ağsı aygıt) olarak adlandırdı. Bu yapının adı, keşfinden 1 yıl sonra Camillo Golgi'nin soyadıyla yeniden adlandırıldı. Ancak bazıları ilk başlarda keşiften şüphe duydu, bu yapının aslında sadece Golgi'nin gözlem tekniğinden kaynaklanan bir optik yanılsama olduğu iddia edildi. 20. yüzyılda modern mikroskopların gelişmesiyle keşif doğrulandı.[3]
Yapısı
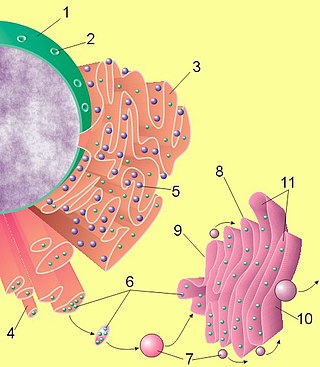
Hem bitkisel hem hayvansal hücrelerde bulunan golgi, sisterna olarak bilinen kıvrımlı zar kümelerinden oluşur. Özellikle bitki hücrelerinde bulunan diktozom adlı (Yunanca, dictyon: ağ + soma: cisim[4]) özgün bir küme bulunur.[5] Bir memeli hücresinde genellikle 40'tan 100'e kadar küme gözlenir.[6] Bir kümede genellikle dört veya sekiz sisterna bulunur.; ancak bazı Protistalarda sisternaların altmışa kadar çıktığı gözlenmiştir.[3] Her sisterna, onu boydan boya geçen kargo proteinlerine yardım eden veya onlara yardım eden özel golgi enzimlerini içeren yassı, etrafı zarlarla çevrili disklerden oluşur.[7]
Sisterna kümesinin dört işlevsel bölgesi vardır: cisim golgi ağı, medial golgi, endo golgi ve trans golgi ağı. Endoplazmik retikulumden veziküler tübüler küme aracılığıyla gelen veziküller, ağ örgüsüyle kaynaşır ve daha sonra paketlenip hedef noktalara gönderildikleri trans golgi ağı boyunca işlenir. Her bölge, bulundukları yere göre içerikleri seçici olarak modifiye eden farklı enzimler içerir.[8] Sisternalar aynı zamanda kendi onarımları için gereken yapısal proteinler taşır.[9]
Görevleri
Hücreler büyük miktarlarda değişik makromolekül sentezler. Golgi aygıtı bu makromoleküllerin hücre sekresyonu[10] (Ekzositoz) veya hücre içi kullanım[11] için paketlenmesinde, türlerine göre ayrılmasında ve modifiye edilmesinde bütünleyici bir göreve sahiptir. Golgi aygıtı öncelikli olarak granüllü endoplazmik retikulumdan gelen proteinleri modifiye eder ve aynı zamanda lipitlerin hücre içinde taşınmasını ve lizozomun oluşumunda görev alır.[11] Bu bakımdan postaneye benzetilebilir; daha sonradan hücrenin çeşitli yerlerine yollayacağı materyalleri paketler ve etiketler.
Sisternada bulunan enzimler karbonhidrat (Glikozilasyon)[12] veya fosfat (Fosforilasyon) ilavesi ile proteinleri modifiye edebilir. Bunu yapabilmek için sitozolden nükleotid şekerler gibi materyalleri içe aktarır. Bu modifikasyonlar aynı zamanda proteinin son varış noktasını belirleyecek olan sinyal sekansını oluşturabilir. Örneğin golgi aygıtı, lizozomlara gidecek olan proteinleri mannoz-6-fosfat ile etiketler
Mitoz esnasında Golgi aygıtı (cisimciği)
Hayvan hücrelerinde, mitozun başlangıcından sonra ayrışır ve kaybolur. Mitozun telofaz evresi esnasında tekrar ortaya çıkar; ancak bunun nasıl olduğu konusunda hala kesin bir kanıya varılamamıştır.[13]
Yanıltıcı bir şekilde bitkilerin veya mayaların hücrelerindeki golgi aygıtlarının hücre döngüsü boyunca bozulmadan durduğu gözlenmiştir. Bu farklılığın nedeni bilinmemekle birlikte kısmen golgi proteinlerinin farkından kaynaklandığı düşünülmektedir.
Ayrıca bakınız
Kaynakça
- ^ Pavelk M, Mironov AA (2008). The Golgi Apparatus: State of the art 110 years after Camillo Golgi's discovery (İngilizce). Berlin: Springer. ISBN 3-211-76309-0.
- ^ a b Fabene PF, Bentivoglio M (Ekim 1998). "1898–1998: Camillo Golgi and "the Golgi": one hundred years of terminological clones". Brain Res. Bull. 47 (3). ss. 195-8. doi:10.1016/S0361-9230(98)00079-3. PMID 9865849.
- ^ a b Davidson MW (13 Aralık 2004). "The Golgi Apparatus". Molecular Expressions. Florida State University. 25 Eylül 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 20 Eylül 2010.
- ^ "Diktozom". Dictionary.com. 9 Eylül 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 20 Eylül 2010.
- ^ Wolfe SA (1993). Molecular and Cellular Biology (İngilizce). Belmont, CA: Wadsworth Pub. Co. s. 828. ISBN 0-534-12408-9.
- ^ Duran JM, Kinseth M, Bossard C, Rose DW, Polishchuk R, Wu CC, Yates J, Zimmerman T, Malhotra V (Haziran 2008). "The role of GRASP55 in Golgi fragmentation and entry of cells into mitosis". Mol. Biol. Cell. 19 (6). ss. 2579-87. doi:10.1091/mbc.E07-10-0998. PMC 2397314 $2. PMID 18385516.
- ^ Becker, Kleinsmith, Hardin, Bertoni (2009). the World of the Cell (İngilizce). San Francisco, CA: Pearson Benjamin Cummings. s. 333, 339. ISBN 0-321-55418-3.
- ^ Krieger M, Scott MP, Matsudaira PT, Lodish HF, Darnell JE. Lawrence Z, Kaiser C, Arnold B (2004). Molecular cell biology (İngilizce) (5th edn bas.). New York: W.H. Freeman and CO. ISBN 0-7167-4366-3.
- ^ "ZFPL1, a novel ring finger protein required for cis-Golgi integrity and efficient ER-to-Golgi transport". 17 Haziran 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Ekim 2011.
- ^ "Regulated Secretion (Golgi): The Movie" (İngilizce). North Dakota State University. 9 Ekim 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Kasım 2010.
- ^ a b Campbell, Neil A (1996). Biology (İngilizce) (4 bas.). Menlo Park, CA: Benjamin/Cummings. s. 122, 123. ISBN 0805319573.
- ^ William G. Flynne (2008). Biotechnology and Bioengineering. Nova Publishers. ss. 45-. ISBN 9781604560671. 29 Haziran 2011 tarihinde kaynağından arşivlendi. Erişim tarihi: 13 Kasım 2010.
- ^ Kimball JW (9 Aralık 2009). "The Golgi Apparatus". Kimball's Biology Pages (İngilizce). 30 Mayıs 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 20 Eylül 2010.
Dış bağlantılar
- Harris E, Cardelli J. "Golgi". Biyoloji Ansiklopedisi (İngilizce). 10 Eylül 2015 tarihinde kaynağından arşivlendi.