Gen aktarım maddesi
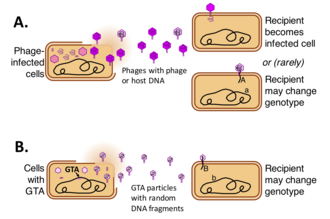
Gen aktarım maddesi (GTA'lar veya GAM), bazı bakteri ve arkeler tarafından üretilen ve yatay gen transferine aracılık eden, DNA içeren virüs benzeri parçacıklardır. Farklı GAM tipleri, çeşitli bakteriyel ve arkeal soylardaki virüslerden bağımsız olarak kaynaklanmıştır. Bu hücreler, hücrede bulunan DNA'nın kısa segmentlerini içeren GAM partikülleri üretir. Parçacıklar üretici hücreden serbest bırakıldıktan sonra, ilgili hücrelere yapışabilir ve DNA'larını sitoplazmaya enjekte edebilirler. DNA daha sonra alıcı hücrelerin genomunun bir parçası olabilir.[1][2][3][4]
Gen aktarım maddesinin keşfi

İlk GAM (GTA) sistemi, 1974 yılında Rhodobacter capsulatus suşlarının karışık kültürlerinde yeni gen birleşimleri ile yüksek bir hücre sıklığına neden olduğu keşfedildi.[5] Sorumlu kalıtımsal özellik taşıyan gen, hücre temasından bağımsız, deoksinükleaza duyarsız ve faj üretimi ile ilişkili olmayan, bilinen gen aktarım mekanizmalarından farklıydı. Varsayılan işlevi nedeniyle gen aktarım maddesi (GTA, şimdi RcGTA) olarak adlandırıldı. Daha yakın zamanda, diğer gen aktarım maddesi sistemleri, filtrelenmiş (hücresiz) kültür ortamının genetik olarak farklı bir tür ile kuluçkalanmasıyla keşfedilmiştir.[3]
GTA (GAM) genleri ve evrimi

Gen Aktarım Maddelerini belirten genler, bir konakçı kromozomuna bütünleşmiş bakteriyofaj (faj) DNA'sından türetilir. Bu tür profajlar genellikle onları kusurlu hale getiren ve faj parçacıkları üretemeyen mutasyonlar edinir. Birçok bakteriyel genom, daha fazla veya daha az kapsamlı silinme ve mutasyon geçirmiş bir veya daha fazla kusurlu profaj içerir. Gen Aktarım Maddesi, özürlü profajlar gibi, profajların mutasyonundan kaynaklanır, ancak faj parçacığının baş ve kuyruk bileşenleri için işlevsel genleri (yapısal genler) ve DNA sarılımı için genleri korurlar. Sistem düzenini ve DNA replikasyonunu belirten faj genleri özgün olarak silinmiştir ve yapısal gen kümesinin ifade edilişi hücresel düzenleyici sistemlerin kontrolü altındadır. GTA üretimine veya alımına katkıda bulunan ilave genler genellikle diğer kromozom bölgelerinde bulunur. Bunlardan bazıları düzenleyici işlevlere sahiptir ve diğerleri doğrudan GTA üretimine (örn. Faj türevi lysis (liziz, hücre çözülmesi) genleri) veya alım ve rekombinasyona (örn. Hücre yüzeyi kapsülü ve DNA taşıma proteinlerinin üretimi) doğrudan katkıda bulunur. Bu GTA ile ilişkili genler genellikle ana GTA gen kümesi ile koordineli düzen içindedir.[6] Fajdan türetilen hücre liziz proteinleri (holin ve endolizin (endolysin)) daha sonra hücre duvarını ve zarını zayıflatır ve zar, hücrenin patlamasına engel olamaz ve GTA (GAM) parçacıklarını serbest bırakır. Her hücre tarafından üretilen GTA parçacıklarının sayısı bilinmemektedir.
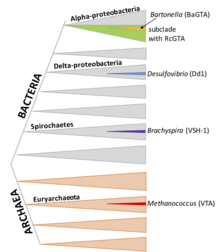
Bazı GTA sistemleri konukçu genomlarına yeni eklenenler gibi görünmektedir, ancak diğerleri milyonlarca yıldır korunmaktadır. Dizi hizalaması çalışmaları yapıldığında (dN / dS analizi), genlerin protein işlevi için doğal seleksiyon ile korunduğunu bildirirler (yani kusurlu uyarlamalar ortadan kaldırılmaktadır).[7][8]
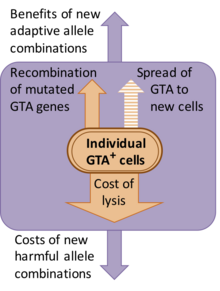
Ancak bu doğal ayıklanmanın niteliği net değildir. Her ne kadar GTA kâşifleri gen transferinin parçacıkların fonksiyonu olduğunu varsaysalar da, gen aktarımının varsayılan yararları nüfus için önemli bir zarara sahiptir. Bu zararın çoğu, GTA üreten hücrelerin GTA parçacıklarını serbest bırakmak için parçalanması gerektiğinden dolayı ortaya çıkar, ancak yeni gen birleşimlerinin oluşmasıyla ilişkili genetik zararlar da vardır, çünkü çoğu yeni birleşim genellikle orijinal birleşimden daha az uygun olacaktır.[9] Alternatif bir açıklama olarak, GTA genlerinin süregelmesidir, çünkü GTA'lar bulaşıcı olarak yeni hücrelere yayılan genetik parazitlerdir. Ancak, bu göz ardı edildi, çünkü GTA parçacıkları karakteristik olarak onları kodlayan genleri içeremeyecek kadar küçüktür. Örneğin, ana RcGTA kümesi 14 kb uzunluğundadır, ancak RcGTA parçacıkları yalnızca 4–5 kb DNA içerebilir.
Çoğu bakteri, GTA'ların varlığı açısından taranmamıştır ve daha birçok GTA sistemi keşfedilmesi beklenilebilir. GTA ile ilgili genler için DNA tabanlı araştırmalar birçok genomda homolog bulmuş olsa da, ancak yorumlama, GTA'ları kodlayan genlerin sıradan profaj genlerinden ayırt edilmesinin zorluğu ile engellenir.[7][8]
GTA üretimi

Laboratuvar kültürlerinde, GTA'ların üretimi özgün olarak GTA genlerinin transkripsiyonunu uyaran özel büyüme koşulları ile maksimuma çıkarılır; GTA'ların çoğu, DNA'ya zarar veren birçok profajı uyaran işlemler tarafından uyarılmaz. Maksimum uyarıcı koşullar altında bile, kültürün sadece küçük bir kısmı GTA üretir, özgün olarak% 1'den az.[10][11]
GTA üretimindeki aşamalar faj enfeksiyonu aşamalarından türetilir. Yapısal genler önce transkripsiyonlanır ve translasyonlanır ve proteinler anlamsız başlara ve serbest kuyruklara birleştirilir. DNA paketleme mekanizmaları daha sonra DNA'yı her başa paketler, baş dolduğunda DNA'yı keser, başa bir kuyruk ekler ve daha sonra yeni oluşturulan DNA ucunu yeni bir boş başa taşır. Profaj genlerinden farklı olarak, GTA'ları kodlayan genlerde genomdan alınma olmaz ve GTA partiküllerinde paketlenmek üzere çoğaltılır. En iyi incelenen iki GTA (RcGTA and BaGTA), hücrede bulunan tüm DNA'ları, GTA kodlayan genlerin fazla temsil edilmemesi için rastgele paketler.[10][12] Her hücre tarafından üretilen GTA partiküllerinin sayısı bilinmemektedir.
Kaynakça
- ^ Lang AS, Westbye AB, Beatty JT (Eylül 2017). "The Distribution, Evolution, and Roles of Gene Transfer Agents in Prokaryotic Genetic Exchange". Annual Review of Virology. 4 (1). ss. 87-104. doi:10.1146/annurev-virology-101416-041624. PMID 28784044.
- ^ Lang AS, Zhaxybayeva O, Beatty JT (Haziran 2012). "Gene transfer agents: phage-like elements of genetic exchange". Nature Reviews. Microbiology. 10 (7). ss. 472-82. doi:10.1038/nrmicro2802. PMC 3626599 $2. PMID 22683880.
- ^ a b Stanton TB (Nisan 2007). "Prophage-like gene transfer agents-novel mechanisms of gene exchange for Methanococcus, Desulfovibrio, Brachyspira, and Rhodobacter species". Anaerobe. 13 (2). ss. 43-9. doi:10.1016/j.anaerobe.2007.03.004. PMID 17513139.
- ^ Grüll MP, Mulligan ME, Lang AS (Ekim 2018). "Small extracellular particles with big potential for horizontal gene transfer: membrane vesicles and gene transfer agents". FEMS Microbiology Letters. 365 (19). doi:10.1093/femsle/fny192
 . PMID 30085064.
. PMID 30085064. - ^ Marrs B (Mart 1974). "Genetic recombination in Rhodopseudomonas capsulata". Proceedings of the National Academy of Sciences of the United States of America. 71 (3): 971-3. doi:10.1073/pnas.71.3.971. PMC 388139 $2. PMID 4522805.
- ^ Westbye AB, Beatty JT, Lang AS (Ağustos 2017). "Guaranteeing a captive audience: coordinated regulation of gene transfer agent (GTA) production and recipient capability by cellular regulators". Current Opinion in Microbiology. 38: 122-129. doi:10.1016/j.mib.2017.05.003. PMID 28599143.
- ^ a b Shakya M, Soucy SM, Zhaxybayeva O (Temmuz 2017). "Insights into origin and evolution of α-proteobacterial gene transfer agents". Virus Evolution. 3 (2): vex036. doi:10.1093/ve/vex036. PMC 5721377 $2. PMID 29250433.
- ^ a b Tamarit D, Neuvonen MM, Engel P, Guy L, Andersson SG (Şubat 2018). "Origin and Evolution of the Bartonella Gene Transfer Agent". Molecular Biology and Evolution. 35 (2): 451-464. doi:10.1093/molbev/msx299 . PMID 29161442.
- ^ Redfield RJ, Soucy SM (2018). "Evolution of Bacterial Gene Transfer Agents". Frontiers in Microbiology (İngilizce). 9: 2527. doi:10.3389/fmicb.2018.02527. PMC 6209664 $2. PMID 30410473.
- ^ a b Hynes AP, Mercer RG, Watton DE, Buckley CB, Lang AS (Temmuz 2012). "DNA packaging bias and differential expression of gene transfer agent genes within a population during production and release of the Rhodobacter capsulatus gene transfer agent, RcGTA". Molecular Microbiology. 85 (2): 314-25. doi:10.1111/j.1365-2958.2012.08113.x. PMID 22640804.
- ^ Kaynak hatası: Geçersiz
<ref>etiketi;Foggetalisimli refler için metin sağlanmadı (Bkz: ) - ^ Kaynak hatası: Geçersiz
<ref>etiketi;Berglund2009isimli refler için metin sağlanmadı (Bkz: )