Ökaryotlar, hücrelerinde bir çekirdek ve –genellikle– organeller içeren bir canlılar grubu olup, bilimsel sınıflandırmada arkeler ve bakterilerle beraber tüm canlıları kapsayan üç ana gruptan biridir.

Eşeysiz üreme tek bir organizmadan yalnızca bu organizmanın genlerini alarak yeni bir canlı üremesidir. Bu üreme yönteminde ploitlik görülmez. Otomiksis durumu dışında eşeysiz üreme sonucu oluşan organizma, ana organizmanın birebir genetik kopyasıdır. Daha dar bir tanımlama gametlerin birleşmesi dışında oluşan üreme yöntemi demek olan agamogenez terimidir. Eşeysiz üreme arkea, bakteri ve protistler gibi tek hücreli organizmalar için ana üreme yoludur. Birçok bitki ve mantar da eşeysiz ürer.

Placozoa, Radiata altâlemine bağlı bir hayvan şubesidir.
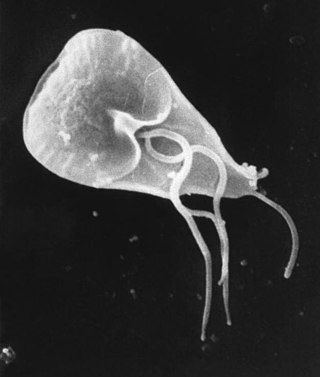
Giardia lamblia. Hexamitidae familyasına ait bir tek protozoon'dur. Giardia intestinalis, Lamblia intestinalis veya Giardia duodenalis olarak da bilinir. Giardiasis veya lambliasis adı verilen hastalığa neden olur.

Crosover veya krossing over ya da parça değişimi mayoz bölünmenin profaz I evresinde görülen, çift halde bulunan kromozomların yaptığı parça değişimine verilen addır. Bunun sonucunda genetik rekombinasyon meydana gelir. Yani farklı kromozomlarda bulunan genlerin alelleri birbiriyle yer değiştirir.

Homolog rekombinasyon, benzer veya aynı dizilere sahip DNA iplikleri arasında nükleotit dizilerinin birbiriyle yer değiştirdiği bir genetik rekombinasyon tipidir. Bu süreç sırasında DNA birkaç kere kesilir, sonra da birleştirilir. Homolog rekombinasyon, DNA'daki çift iplikli kırıkların hatasız tamirinde kullanılmanın yanı sıra, mayoz sırasında krosover yoluyla yeni DNA dizi bileşimlerinin (kombinezonlarının) oluşumunu da sağlar. DNA'daki yeni bileşimler genetik varyasyonlar oluşturur. Genetik varyasyonlar yeni, bir olasılıkla yararlı olabilecek alel kombinasyonlarıdır, bunların üreyen canlı topluluklarda oluşmaları, bu değişiklikleri taşıyan bireylerin değişen çevresel şartlara evrimsel adaptasyon göstermelerini sağlar.

RAD51, bir insan genidir, bu genin ürünü olan ve aynı isme sahip olan protein DNA çift iplikçikli kesiklerini tamir eder. Bu protein RAD51 protein ailesinin üyelerinden biridir. RAD51 protein ailesi üyeleri bakteriyel RecA ve maya Rad51 ile homologdur. Protein, mayadan insana kadar çoğu ökaryotta korunmuştur.
Bir minisatelit, DNA içinde tekrar eden 10-60 bazlık bir dizidir. Bunlar insan genomunda binden çok konumda bulunur. Bazı minisatelitlerde DNA ipliklerinden biri pürin öbürü pirimidin ağırlıklı olur, bazıları ise “GGGCAGGANG” bazlarından oluşan bir merkez dizi içerirler. Bu dizinin kromozomlar arasında dizi takasına yol açtığı öne sürülmüştür. Başka bir görüşe göre ise, minisatelitlerde kopya sayısı varyasyonunun nedeni, yakınında bir çift iplik DNA kırılma noktasının bulunmasıdır. DNA ikileşmesi sırasındaki sorunlar, örneğin DNA ipliklerinden birinin öbürüne göre kayması sonucu hatalar olmakta ve minisatelitin tekrarlayan dizi sayısında bir değişiklik meydana gelmektedir. En çok değişkenlik gösteren lokus CEB1 (D2S90)'dir.

Birçok türün canlıları her biri cinsiyet veya eşey olarak bilinen dişi ve eril çeşitlerine özelleşmiştir. Eşeyli üreme genetik özelliklerin karışımı ve birleşimini içerir: Gamet olarak bilinen özelleşmiş bu hücreler her bir ebeveynden özelliklerini kalıtsal yolla alan yavrular oluşturmak için birleşir. Gametler form ve fonksiyonda özdeş (izogami) olabilir, fakat birçok durumda iki cinsiyete özgü gamet (heterogamet) tiplerinin oluşması (anizogami) gibi bir asimetri evrilebilir. Eril gametleri küçüktür, hareket edebilirler ve genetik bilgilerini bir mesafe boyunca taşıyabilmeye uyum sağlamışlardır; dişi gametleri ise büyüktür, hareket edemezler ve genç canlının ilk gelişimi için gerekli olan besini barındırırlar. İnsanlar ve diğer memeliler arasında, eriller genellikle XY kromozomlarını taşırken, dişiler ise genellikle XX kromozomlarını taşırlar ve XY cinsiyet belirleme sisteminin bir parçasıdır.

Archaeplastida, kırmızı algler (Rhodophyta), yeşil algler ve kara bitkilerini ve glokofitler gibi bazı küçük grupları içeren önemli bir ökaryot grubudur. Archaeplastida'nın tüm soyları, fotosentetik olmayan ökaryotrofik bir kamçılı olan Rhodophyta'nın kardeş taksonu Rhodelphidia dışında fotoototrofiktir. Archaeplastida üyeleri, iki zarla çevrili kloroplastlara sahiptir, bu özellikleri bu canlıların bir siyanobakteri ile beslenerek endosimbiyoz olayı yoluyla doğrudan elde edildiğini düşündürür. Amoeboid Paulinella chromatophora'nın yanı sıra kloroplastlara sahip diğer tüm gruplar, ikincil olarak kırmızı veya yeşil alglerden elde edildiklerini düşündüren üç veya dört zarla çevrili kloroplastlara sahiptir. Kırmızı ve yeşil alglerin aksine, glokofitler hiçbir zaman ikincil endosimbiyoz olaylarına dahil olmamıştır.

Amorphea veya Unikonta, Amoebozoa ve Obazoa'yı içeren taksonomik bir grupdur. Bu sınıfın üyelerinin taksonomik yakınlıkları ilk olarak 2002 yılında Thomas Cavalier-Smith tarafından açıklanmıştır.

Opisthokontlar, hem hayvan hem de mantar alemlerini içeren geniş bir ökaryot grubudur. Daha önce "Fungi / Metazoa grubu" olarak adlandırılan opisthokontlar, genellikle bir sınıf olarak tanınır. Opisthokontlar, Apusomonadida ve Breviata ile birlikte daha büyük Obazoa kladını oluşturur.

Viridiplantae, yaklaşık 450.000-500.000 tür içeren ve hem karasal hem de sucul ekosistemlerde önemli roller oynayan ökaryotik canlılar grubudur. Öncelikle sucul olan yeşil algler ve içlerinden çıkan kara bitkilerinden oluşurlar. Yeşil algler, geleneksel sınıflandırmada kara bitkilerini içermez ve bu da yeşil algleri parafiletik bir grup yapar. Kara bitkilerinin yeşil alglerin içinden çıktığının anlaşılmasından bu yana, bazı yazarlar bitkileri de yeşil alglere atıyorlar. Hücre duvarlarında selüloz bulunan hücrelere ve klorofil a ve b içeren ve fikobilin içermeyen siyanobakterilerle endosimbiyozdan türetilen birincil kloroplastlara sahiptirler.

Holozoa, hayvanları ve en yakın tek hücreli akrabalarını içeren ancak mantarları hariç tutan bir canlı grubudur. Holozoa ayrıca tunik cinsi Distaplia için kullanılan eski bir isimdir.
Filozoa, Opisthokonta içindeki monofiletik bir gruptur. Bunlar hayvanları ve onların en yakın tek hücreli akrabalarını içerir.

Holomycota veya Nucletmycea, Holozoa'nın kardeş grubu olan temel bir Opisthokont klanıdır. Cristidiscoidea ve mantar (fungi) aleminden oluşur. Holomycota'nın en eski soyu olarak nükleeriidlerin, tek hücreli serbest yaşayan fagotrofik amiplerin konumu, hayvanların ve mantarların bağımsız olarak ortak bir tek hücreli atadan karmaşık çok hücreliliği edindiğini ve ozmotrofik yaşam tarzının daha sonra ortaya çıktığını göstermektedir. bu ökaryotik soyun ayrışmasında. Opisthosporidians, aphelidleri, Microsporidia ve Cryptomycota, üç endoparazit grubunu içeren yakın zamanda önerilen bir taksonomik gruptur.

SAR veya Harosa; Stramenopiles, Alveolata ve Rhizaria'dan oluşan ve Bikonta kladına bağlı bir kladdır. SAR ismi Stramenopiles, Alveolata, Rhizarianın kısaltmasıdır.

Tek hücreli organizma olarak da bilinen tek hücreli canlı, birden fazla hücreden oluşan çok hücreli bir organizmanın aksine, tek bir hücreden oluşan bir organizmadır. Organizmalar iki genel kategoriye ayrılır: prokaryotik organizmalar ve ökaryotik organizmalar. Prokaryotların çoğu tek hücrelidir ve bakteri ve arkea olarak sınıflandırılır. Birçok ökaryot çok hücrelidir, ancak protozoa, tek hücreli algler ve tek hücreli mantarlar gibi bazıları tek hücrelidir. Tek hücreli organizmaların yaşamın en eski formu olduğu düşünülmektedir; ilk protoseller muhtemelen 3.8-4.0 milyar yıl önce ortaya çıkmıştır.

Excavata, Ökaryot üst alemine ait eski bir tek hücreli organizma üst grubudur. İlk olarak 1999'da Simpson ve Patterson tarafından önerilmiş ve 2002'de Thomas Cavalier-Smith tarafından resmi bir takson olarak tanıtılmıştır. Çeşitli serbest yaşayan ve simbiyotik formlar içerir ve Giardia ve Trichomonas gibi bazı önemli insan parazitlerini de içinde barındırır. Daha önce artık modası geçmiş Protista krallığına dahil olduğu düşünülüyordu. Kamçılı yapılarına göre sınıflandırıldılar ve en temel kamçılı soy olarak kabul edildiler.

Spermatositler hayvanlarda bulunan erkek üreme hücreleridir ve olgunlaşmamış üreme hücreleri olan spermatogonyumdan türetilirler. Spermatositler testislerde, özellikle de seminifer tübül adı verilen bir yapı içerisinde bulunur. Primer ve sekonder spermatositler olmak üzere iki tip spermatosit vardır. Birincil ve ikincil spermatositler spermatositogenez ile oluşur.