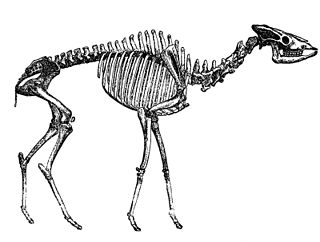
Daeodon
| Daeodon | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Daeodon shoshonensis'in kafatası, Carnegie Museum of Natural History | |||||||||||||
| Biyolojik sınıflandırma | |||||||||||||
| |||||||||||||
| Tip tür | |||||||||||||
| †Daeodon shoshonensis Cope, 1878 | |||||||||||||
| Türler | |||||||||||||
| Sinonimler | |||||||||||||


Daeodon, geç Oligosen ve en erken Miyosen sırasında Kuzey Amerika'da yaşayan soyu tükenmiş bir entelodont çift toynaklı cinsidir. Tip türü, Cope tarafından çok şüpheli bir holotip tarafından tanımlanan Daeodon shoshonensis'tir. Bazı yazarlar onu Dinohyus hollandi ve diğer birkaç türle eş anlamlı olarak kullanırlar (metne bakınız), ancak tanı materyali eksikliğinden dolayı bu en iyi ihtimalle şüphelidir.
Bu ailenin Daeodon'a benzeyen bir diğer büyük üyesi, Asyalı Paraentelodon'dur, ancak çok eksik malzeme ile bilinir.[1][2]
Etimoloji
Cope'un orijinal açıklamasında belirtilmese de, Daeodon adı Yunanca "düşman" veya "korkunç" anlamına gelen daios ve "dişler" anlamına gelen odon kelimelerinden gelmektedir.[3]
Sınıflandırma
Daeodon cinsi, 1878'de Amerikalı anatomist ve paleontolog Edward Drinker Cope tarafından tanımlandı. Cope, onu bir tek toynaklı olarak sınıflandırdı ve Menodus ile yakından ilişkili olduğu düşünülüyordu.[4] Bu sınıflandırma, Peterson (1909) tarafından derhal bir Daeodon türü olarak atanan, aynı kayalardan çok benzer ve çok daha eksiksiz bir hayvan olan 1905'te " Elotherium calkinsi" nin[5] tanımına kadar devam etti.[6] Bu, Daeodon'un Entelodontidae ailesinin bir üyesi olarak yeniden sınıflandırılmasına yol açtı. Daeodon ve diğer entelodontlar arasındaki kesin ilişkiler iyi anlaşılmamıştır, bazı yazarlar (Lucas ve diğerleri, 1998), Daeodon'un Archaeotherium gibi daha önceki Kuzey Amerika entelodontlarından ziyade Paraentelodon'a daha büyük morfolojik benzerliğini, Büyük Asya entelodontlarının Geç Oligosen'deki Kuzey Amerika'ya göçen soyundan geldiğine dair kanıt olarak düşünmektedir.[7] Bununla birlikte, hem Paraentelodon hem de Daeodon'da bulunanları anımsatan özellikler gösteren farklı Archaeotherium örneklerinin varlığı, her iki cinsin de aslında Kuzey Amerika'daki bir ortak atadan gelme olasılığını artırmaktadır.[1][8]
Türler
Daeodon'un türü, Oregon'daki John Day Formasyonu'ndan bir alt çene parçasına dayanan D. shohonensis'tir. Sonraki yıllarda D. calkinsi, D. mento[9] ve D. minor gibi birkaç başka tür de cinse atanmıştır.[10] 1945'ten beri, diğer iki taksonun aslında Daeodon'un küçük eşanlamlıları olduğu öne sürülmüştü,[11] ancak bu başvurunun resmileştirilmesi Lucas ve arkadaşlarının (1998) çalışmasına kadar gerçekleşmedi.[7] Cope'un rakibi O. Charles Marsh tarafından adlandırılan ve Dinohyus hollandi ve Ammodon leidyanum'un,[12] Nebraska'daki Agate Springs ocağından alınan tam bir iskeleti,[13] birbirinden ayırt edilemez bulundu ve sırayla her ikisi de D. shoshonensis'ten ayırt edilemezdi.[7] Daeodon'dan geçici olarak hariç tutulan D. calkinsi dışında, daha önce tanınan Daeodon türleri de D. shoshonensi ile eşanlamlı hale getirildi.[7] Aynı yıl, belirsiz bir entelodont, Boochoerus humerosum, da Foss ve Fremd (1998) tarafından Daeodon ile eşanlamlı hale getirildi ve ayrı bir tür olarak statüsü korunsa da, farklılıkların hala bireysel veya popülasyon varyasyonundan veya cinsel dimorfizm'den kaynaklanma ihtimali vardır.[14]
Açıklama
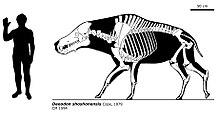
Daeodon shoshonensis bilinen en büyük entelodonttur;[7] bilinen yetişkin bireylerin kafatasları yaklaşık 90 cm uzunluğundaydı ve omuzda yaklaşık 1,77 metre boyundaydı.[6] Diğer entelodontlardan benzersiz diş özellikleri, kafatasının elmacık kemiği flanşlarının şekli, Archaeotherium'a kıyasla nispeten küçük boyutu, çene tüberkülünün küçük boyutu ve ayrıca alt bacak kemiklerinin, füzyonu, sazan ve tarsusunun özellikleriyle ayırt edilir.[1][6][15] Diğer entelodontlar gibi, bacakları uzun ve inceydi, ön bacak kemikleri birbirine kaynaşmıştı[7][15] ve her ayağında sadece iki toynak vardı.[6][7] Aynı zamanda, ağırlığı çoğunlukla torasik omurların uzun dikenlerine bağlı kaslar ve tendonlar tarafından desteklenen, günümüz bizon ve beyaz gergedanlarına benzer şekilde, başının boyutuna göre nispeten hafif yapılı bir boynu vardı.[15]
Paleoekoloji
Doğal ortam
Daeodon, Kuzey Amerika'da geniş bir yelpazeye sahipti ve Akik Fosil Yataklarında bulunan birçok fosil, yoğun ormanlar ve geniş çayırlar arasındaki bir geçiş döneminde bir ortamı temsil ediyor, bu, muhtemelen orta Miyosen'de yok olmalarının önemli bir nedeniydi.[16] Archaeotherium gibi daha fazla bazal entelodontdan daha fazla yüzeysel bir vücut planıyla otlaklara uyum sağladı, çiğ pençelerini tamamen kaybetti, proksimal olarak kaynaşmış metacarpallara ve bizona benzer omuz kaslarına sahipti.[15][17]
Diyet

Daeodon diğer tüm entelodontlar gibi omnivordur. Emaye desenleri, et ve kemiklerin yanı sıra fındık, kök ve asma yemeyi önerir. Pekariler, su aygırları ve ayılarla olan yüzeysel benzerlik, Daeodon'un hangi bitkileri yemiş olabileceği konusunda geniş bir yelpazeye işaret ediyor. O zamanlar Kuzey Amerika'nın kurak mevsimleri çok sertleşebilirdi, bu yüzden su alımlarını üzüm asmaları yiyerek desteklemiş olabilirler. Etçilliğinin kapsamı tartışılır, ancak diş aşınması, kemik kırma ve et parçalama konusunda uzmanlaştıklarını ve chalicothere kemiklerindeki ısırık izleri, büyük otçulları avladıklarını veya temizlediklerini düşündürür. Foss (2001), kafasının büyük bir avı alt etmek için etkili olamayacak kadar ağır olduğunu, bu nedenle yalnızca çöpçülüğe dayanmış olması gerektiğini, ancak koşmak için bizon benzeri adaptasyonlarının, yırtıcıların stereoskopik görme özelliğinin ve entelodontlarda yırtıcılığın kanıtlarının olduğunu savunuyor.[18] Diyetlerinin belirsizliği, duruma bağlı olarak ihtiyaç duydukları her şeyi yiyen ayılara benzer fırsatçı omnivorlar olduklarını gösteriyor.[19]
Davranış
Entelodontlar, kafataslarındaki diş izlerinden bilinen tür içi yüz ısırmasına katıldılar. Erkekler, kas ekleri olarak işlevlerine ek olarak, muhtemelen mandibular tüberküllerini koruma olarak kullanarak, baskınlık için savaşırlardı.[19] Jugal korumaların cinsel dimorfizmi Archaeotherium'da mevcuttur ve daha küçük bir Daeodon numune boyutuyla, Daeodon için bu tür bir dimorfizm göz ardı edilemez. Dimorfikse, genişletilmiş jugallerin işlevi, kimyasal iletişim için sahip olunan orman domuzlarına benzer büyük yörünge öncesi bezleri destekleyen, muhtemelen sergilenecekti.
Ayrıca bakınız
Kaynakça
- ^ a b c The Evolution of Artiodactyls. JHU Press. 2007. ISBN 9780801887352. 12 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ Бернарская фауна олигоценовых позвоночных (The Benarskaya Fauna of Oligocene Vertebrates). Metsniereba, Tbilisi. 1964. s. 109-133. 12 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 26 Eylül 2020.
- ^ Composition of scientific words: A manual of methods and a lexicon of materials for the practice of logotechnics. Smithsonian Institution Press. 1954. ISBN 978-0874740011.
- ^ Cope (1878). "On some characters of the Miocene fauna of Oregon". Paleontological Bulletin. 30: 1-16.
- ^ Sinclair (1905). "New and imperfectly known rodents and ungulates from the John Day Series". Bull. Dept. Geology, Univ. California. 4: 132-134.
- ^ a b c d Peterson (1909). "A revision of the Entelodontidae". Memoirs of the Carnegie Museum. 4 (3): 41-158. 16 Ekim 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ a b c d e f g Lucas (1998). "Taxonomy and distribution of Daeodon, an Oligocene-Miocene entelodont (Mammalia: Artiodactyla) from North America". Proceedings of the Biological Society of Washington. 111 (2): 425-435. 16 Ekim 2018 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ Foss (2001). "Biostratigraphy of the Entelodontidae (Mammalia: Artiodactyla) from the John Day Basin, Oregon". Paleobios. 21: 53.
- ^ Allen (1926). "Fossil mammals from South Carolina". Bulletin of the Museum of Comparative Zoology. 67: 447-467.
- ^ Loomis (1932). "Two new Miocene entelodonts". Journal of Mammalogy. 13 (4): 358-362. doi:10.2307/1374141.
- ^ Simpon (1945). "The principles of classification and a classification of mammals". Bulletin of the American Museum of Natural History. 85: 1-350.
- ^ Peterson (1905b). "A correction of the generic name (Dinochoerus) given to certain fossil remains from the Loup Fork Miocene of Nebraska". Science. 22 (570): 719. doi:10.1126/science.22.570.719. PMID 17729479. 12 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ Peterson (1905a). "Preliminary note on a gigantic mammal from the Loup Fork Beds of Nebraska". Science. 22 (555): 211-212. doi:10.1126/science.22.555.211. PMID 17835750. 12 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ Foss (1998). "A survey of the species of Entelodonts (Mammalia, Artiodactyla) of the John Day Basin, Oregon". Dakoterra. 5: 63-72.
- ^ a b c d "Entelodontidae". Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Cambridge University Press. 1998. ISBN 9780521355193. 12 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ "Arşivlenmiş kopya" (PDF). 17 Ocak 2019 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 12 Ağustos 2021.
- ^ "Arşivlenmiş kopya". 12 Ağustos 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ "Abstract of Papers. Fifty-ninth Annual Meeting Society of Vertebrate Paleontology". Journal of Vertebrate Paleontology. 19 (3): A1-A93. 1999. ISSN 0272-4634. 13 Temmuz 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 12 Ağustos 2021.
- ^ a b Foss, S. E., 2001, Systematics and paleobiology of the Entelodontidae (Mammalia, Artiodactyla) [Ph.D. dissertation]: Dekalb, Northern Illinois University
Konuyla ilgili yayınlar
- Russell, Loris S. (1980). Tertiary mammals of Saskatchewan, Part V : the Oligocene Entelodonts. Toronto: Royal Ontario Museum. ISBN 0-88854-254-2. OCLC 7437855.
Dış Bağlantılar
Videolar
- Prehistoric Predators - Hell Pig (41:34)