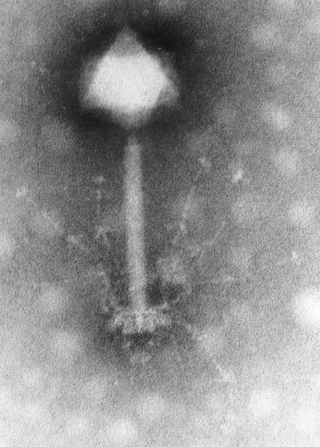
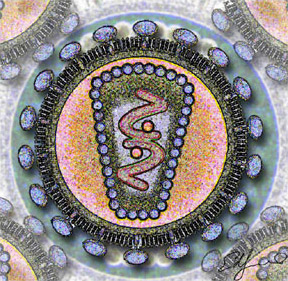
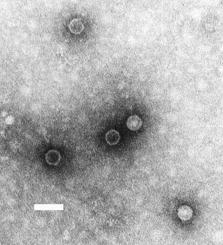
DNA virüsleri
Genetik madde olarak DNA bulunduran ve replikasyon için DNA'ya bağımlı DNA polimeraz enzimine gereksinim duyan virüslerdir. Nükleik asitleri genellikle çift iplikçikli DNA yapısındadır (çiDNA) ancak tek iplikçikli DNA yapısında olanlar da mevcuttur (tiDNA). DNA virüsleri Baltimor sınıflandırma sisteminde I. Grup ya da II. Grup üyeleridir. Tek iplikçikli DNA'lar genellikle enfekte hücrede çift iplikçikli yapıya dönüştürülür. Hepatit B virüsünü de içeren VII. Grup virüsleri DNA genomuna sahip olmalarına rağmen Baltimor sınıflandırmasında DNA virüslerinden ayrı bir sınıfta yer almışlardır, bunun nedeni bu virüslerin ters transkripsiyon yapan virüs olması ve RNA aracılı replikasyon yapmasıdır. DNA virüsleri çiçek hastalığı, herpesvirüs hastalıkları ve su çiçeği gibi önemli hastalıklara neden olabilmektedirler.
Grup I: çiDNA virüsleri
Bu grubun genom çeşitliliği önemli farklılılar gösterir. Bazıları çembersel (sirküler) genoma (Baculoviridae, Papovaviridae ve Polydnaviridae) sahip iken, bazıları da çizgisel (lineer) genoma sahiptirler (Adenoviridae, Herpesviridae ve bazı fajlar). Bazı aileler çembersel ve çizgisel arasında değişim gösterebilen genoma sahip iken (T4 fajı ve bazı Iridoviridaeler). Bazıları da kovalent olarak kapalı uçlu çizgisel genoma sahiptirler (Poxviridae ve Phycodnaviridae).
Konak aralığı
Kaudovirüs takımı türleri,Kortikovirüs ve Tetivirüs aileleri bakterileri enfekte ederler.
Ligamenvirüs takımı türleri, Ampullavirüs, Bicaudavirüs, Clavavirü, Fusellovirü, Globulovirüs, Guttavirüs ve Turrivirüs aileleri; hipertermofilik arkelerin Crenarchaeota türlerini enfekte ederler.
Herpesvirales takımı türleri, Adenovirüs, Asfarvirüs, Iridovirüs, Papillomavirüs, Polyomavirüs ve Poxvirüs aileleri; omurgalıları enfekte ederler..
Ascovirus, Baculovirus, Hytrosaviridae, Iridoviridae, Polydnaviruses aileleri ve Nudivirus cinsi; böcekleri enfekte ederler.
Mimivirüs ailesi türleri, Marseillevirüs, Megavirüs, Mavirüs virofajı ve Uydu virofajı türleri protozoaları enfekte ederler
Nimaviridae ailesi türleri kabukluları enfeke ederler.
Phycodnavirüs ailesi türleri ve Organik Göl virofajı türleri algleri enfekte ederler. Bunlar bitkileri enfekte eden bilinen tek çiDNA virüsleridir.
Plasmavirüs ailesi türleri; Mollicutes sınıfı türlerini enfekte eder.
Pandoravirüs ailesi türleri amipleri enfekte eder.
Dinodnavirus cinsi türleri ateşrengi algleri enfekte ederler.
Rhizidiovirus cinsi türleri stramenopilleri enfekte edeler.
Salterprovirus ve Sphaerolipoviridae cinsi türleri euryarchaeota cinsine enfekte ederler.
Taksonomi
- Takım Caudovirales
- Aile Myoviridae—Enterobacteria phage T4 dahil
- Aile Podoviridae—Enterobacteria phage T7 dahil
- Aile Siphoviridae—Enterobacteria phage λ dahil
- Takım Herpesvirales
- Aile Alloherpesviridae
- Aile Herpesviridae— insan herpesvirüsleri ve Varicella Zoster virus dahil
- Aile Malacoherpesviridae
- Takım Ligamenvirales
- Aile Lipothrixviridae
- Aile Rudiviridae
- Belirlenmemiş aileler
- Aile Adenoviridae—insanlarda adenovirüs enfeksiyonu yapanlar dahil.
- Aile Ampullaviridae
- Aile Ascoviridae
- Aile Asfarviridae—Afrika domuz ateşi virüsü dahil
- Aile Baculoviridae
- Aile Bicaudaviridae
- Aile Clavaviridae
- Aile Corticoviridae
- Aile Fuselloviridae
- Aile Globuloviridae
- Aile Guttaviridae
- Aile Hytrosaviridae
- Aile Iridoviridae
- Aile Marseilleviridae
- Aile Mimiviridae
- Aile Nudiviridae
- Aile Nimaviridae
- Aile Pandoraviridae
- Aile Papillomaviridae
- Aile Phycodnaviridae
- Aile Plasmaviridae
- Aile Polydnaviruses
- Aile Polyomaviridae—Simian virüs 40, JC virüs, BK virüs dahil
- Aile Poxviridae— Sığır çiçeği virüsü, çiçek hastalığı virüsü dahil.
- Aile Sphaerolipoviridae
- Aile Tectiviridae
- Aile Turriviridae
- Belirlenmemiş cinsler
- Dinodnavirus
- Salterprovirus
- Rhizidiovirus
- Belirlenmemiş türler
- Abalone shriveling syndrome-associated virus
- Bandicoot papillomatosis carcinomatosis virus
- KIs-V
- Lentille virus
- Megavirus
- Mollivirus sibericum virus
- Phaeocystis globosa virus
- Pithovirus
- Virofajlar
- Ace Lake Mavirus virophage
- Dishui Lake virophage 1
- Mavirus virophage
- Organic Lake virophage
- Phaeocystis globosa virus virophage
- Rio Negro virophage
- Sputnik virophage
- Sputnik virophage 2
- Yellowstone Lake virophage 1
- Yellowstone Lake virophage 2
- Yellowstone Lake virophage 3
- Yellowstone Lake virophage 4
- Yellowstone Lake virophage 5
- Yellowstone Lake virophage 6
- Yellowstone Lake virophage 7
- Zamilon virophage
- Zamilon virophage 2
Pleolipovirüsler
Pleolipovirüsler olarak bilinen bir gruptur, benzer genom organizasyonuna sahip olmalarına rağmen, tek ya da çift iplikçikli DNA genomu yapısında olmalarıyla farklılıklar gösterirler.[1][2] Bu grup mevcut sınıflandırma sistemine uymamaktadır ve bu grup için yeni bir takson gerekmektedir.
Bu virüsler litik değillerdir ve virionları lipit veziküllerin kapladığı genomlar ile karaktrizedir.[3] Nükleoproteinlere sahip değillerdir. Viral zardaki lipitler, konak hücre zarından seçici olmayan bir şekilde elde edilmiştir. Virionlar iki ya da üç adet büyük yapısal protein barındırırlar.
Bu grupta aşağıdaki virüsler bulunmaktadır:
- Haloarcula hispanica pleomorphic virus 1
- Halogeometricum pleomorphic virus 1
- Halorubrum pleomorphic virus 1
- Halorubrum pleomorphic virus 2
- Halorubrum pleomorphic virus 3
- Halorubrum pleomorphic virus 6
- SNJ2
Grup II: tiDNA virüsleri

Bakteriyofajların ilk olarak 1927'de keşfedilmelerine rağmen, sadece 1959'da Sinshemer çalışmasında Phi X 174 fajının tek iplikçikli DNA yapısında olabileceğini öne sürmiştür.[4][4] Bu keşfe kadar DNA virüslerinin birçoğunun göreceli olarak çift iplikçikli yapıda olduğu düşünülmekteydi.[5][6][7]
Taksonomi
Aileler genom doğası ve konak aralığı temel alınarak gruplandırılmışlardır. Aşağıda bu aileler yer almaktadırlar.
- Aile Anelloviridae
- Aile Bacillariodnaviridae
- Aile Bidnaviridae
- Aile Circoviridae
- Aile Geminiviridae
- Aile Inoviridae
- Aile Microviridae
- Aile Nanoviridae
- Aile Parvoviridae
- Aile Spiraviridae
Sınıflandırma
Çembersel tek iplikçikli virüslerin bir bölümünün dört tipe ayrıldığı öne sürülmüştür.[8] Bu bölümlendirmenin, onların filogenetik ilişkilerini yansıttığı görülmektedir.
Tip I, küçük çembersel DNA (yaklaşık 2-kb) ile karakterizedir, Rep proteini ve büyük açık okuma alanı (ORF) zıt yönlerdedir. Bu tip: sirkovirüslerin, geminivirüslerin ve nanovirüslerin karakteridtikleridir.
Tip II, genomları benzersiz ve ayrı iki Rep ve ORFlardan oluşur.
Tip III, genomları aynı yönelimde iki büyük ORF içerir. Bu tipik bit anellovirüs düzenidir.
Tip IV, genomları 8'e kadar olabilen ORF ile beraber 4-kb'ı bulan büyük genomlardandır. Bu genom tipi İnovirüsler ve Mikrovirüslerde bulunur.
Konak aralığı
Bidnavirüs ve Parvovirüs aileleri çizgisel genoma sahip iken diğer aileler çembersel genoma sahiptirler Bidnavirüsler iki parçalı genoma sahiptirler ve omurgalıları enfekte ederler. İnovirüsler ve Mikrovirüsler bakterileri enfekte ederler. Anellovirüsler ve Sircovirüsler hayvanları (memelileri ve kuşları ayrı ayrı olarak) enfekte ederler; ve Geminivirüsler ve Nanovirüsler bitkileri enfekte ederler. Bacillariodnavirüsler diatomları enfekte ederler ve eşsiz genoma sahiptirler: ana kromozom daireseldir (~6 kilobaz uzunluğunda): küçük kromozom doğrusaldır (~1 kilobaz uzunluğunda) ve ana kromozomun tamamlayıcısıdır. Spiravirüsler üyeleri arkeleri enfekte ederler. Mycodnavirüsler üyeleri mantarları enfekte ederler.
Moleküler biyoloji
Bu gruptaki virüsler, genom replikasyonu için çift iplikçikli DNA aracısı replikatif bir forma ihtiyaçları vardır. Bu form, normal olarak konak hücre DNA polimerazı yardımıyla virüs DNA'sından oluşturulur.
Kaynakça
- ^ Pietilä MK, Atanasova NS, Manole V, Liljeroos L, Butcher SJ, Oksanen HM, Bamford DH (2012).
- ^ Sencilo A, Paulin L, Kellner S, Helm M, Roine E (2012). "Related haloarchaeal pleomorphic viruses contain different genome types". Nucleic Acids Res. 40 (12). ss. 5523-34. doi:10.1093/nar/gks215. PMC 3384331 $2. PMID 22396526.
- ^ Pietilä MK, Atanasova NS, Manole V, Liljeroos L, Butcher SJ, Oksanen HM, Bamford DH (2012). "Virion architecture unifies globally distributed pleolipoviruses infecting halophilic archaea". J Virol. 86 (9). ss. 5067-79. doi:10.1128/JVI.06915-11. PMC 3347350 $2. PMID 22357279.
- ^ a b Sinshemer RL (1959).
- ^ Desnues C, Rodriguez-Brito B, Rayhawk S, Kelley S, Tran T, Haynes M, Liu H, Furlan M, Wegley L, Chau B, Ruan Y, Hall D, Angly FE, Edwards RA, Li L, Thurber RV, Reid RP, Siefert J, Souza V, Valentine DL, Swan BK, Breitbart M, Rohwer F (2008).
- ^ Angly FE, Felts B, Breitbart M, Salamon P, Edwards RA, Carlson C, Chan AM, Haynes M, Kelley S, Liu H, Mahaffy JM, Mueller JE, Nulton J, Olson R, Parsons R, Rayhawk S, Suttle CA, Rohwer F (2006).
- ^ Roux S, Krupovic M, Poulet A, Debroas D, Enault F (2012). "Evolution and diversity of the Microviridae viral family through a collection of 81 new complete genomes assembled from virome reads". PLoS ONE. 7 (7). ss. e40418. doi:10.1371/journal.pone.0040418. PMC 3394797 $2. PMID 22808158.
- ^ Rosario K, Duffy S, Breitbart M (2009).
Konuyla ilgili yayınlar
- Fauquent, C. M.; Mayo, M. A.; Maniloff, J.; Desselberger, U.; Ball, L. A. (27 Mayıs 2005). "Uluslararası Virüs Taksonomisi Komitesinin 8. Raporu". Academic Press. US National Center for Biotechnology Information, National Library for Medicine, National Institutes of Health. 6 Kasım 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 9 Ağustos 2016.Academic Press. Birleşik Devletler Ulusal Biyoteknoloji Merkezi, Ulusal Sağlık Enstitüsü, Ulusal Tıp Kütüpanesi.