DNA nanoteknolojisi
DNA nanoteknolojisi nanoteknolojinin bir alt sahasıdır, DNA ve diğer nükleik asitlerin moleküler tanıma özelliklerini kullanarak yeni moleküler yapılar oluşturmayı amaçlar. Bu sahada, DNA kalıtsal bilgi taşıyıcısı olarak değil, yapısal bir malzeme olarak kullanılır. Bunun uygulaması moleküler özbirleşme (İngilizce self assembly) ve DNA hesaplamasıdır.
Tarih

DNA nanoteknoloji kavramı 1980'li yılların başlarında Nadrian Seeman tarafından icat edilmiştir.[1] Bir kristalograf olan Seeman, bazı moleküllerin kristalleştirilmesini sağlayacak şartların bulunmasının tahmine ve tesadüflere dayalı olmasından yılmıştı. 1980'de, M. C. Escher'in Derinlik adlı gravürü ona ilham vermiş, üç boyutlu bir DNA örgüsünün (latisin), kristallenmesi amaçlanan molekülleri yönledirebileceğini fark ettirmiştir.[2][3] 1991'de Seeman'ın laboratuvarı, DNA'dan oluşan bir kübün sentezi hakkındaki raporunu yayımladı. Seeman, nano-ölçekli bu ilk üç boyutlu cisim için 1995'te Feynman Nanoteknoloji Ödülünü almıştır. Nano-kübün ardından, DNA'dan yapılmış kesik oktahedron gelmiş ama anlaşılmıştır ki bu cisimler üç boyutlu bir örgü oluşturabilecek kadar rijit değildir.[1][2]
Seeman daha rijit olan "DX" motifini geliştirmiş ve 1988'de Erik Winfree ile birlikte iki boyutlu DX "karo"larından oluşan iki boyutlu örgülerin oluşturulmasını yayımlamıştır. Bu karolara dayalı yapıların bir diğer avantajı, DNA hesaplamasının gerşekleştirilme olanağını sağlamalarıydı. Winfree ve Paul Rothemund 2004'te bunu gösterdiler ve bunun için 2006'da Feynman Nanoteknoloji Ödülü'nü paylaştılar.[1][2]
Bu saha dallanmaya devam etmektedir. İlk DNA makinası (bir girdinin etkisiyle yapısını değiştiren bir yapısal motif) 1999'da gösterilmiştir. Nanomimari 1987'de Seeman tarafından önerilmiştir ve 2006'da bu sahanın ilk uygulamaları gösterilmiştir. Gene 2006'da Rothemund ilk DNA origami tekniği ile, herhangi bir şekle sahip katlanmış DNA moleküllerinin kolaylıkla oluşturulabileceğini göstermiştir. 2009'da Seeman üç boyutlu bir örgü (kafes) sentezini yayımlamıştır, bu amaçta çalışmaya başlamasından neredeyse otuz yıl sonra.
Temel kavramlar
DNA nanoteknolojisinde dallı DNA yapıları kullanılınca faydalı özellikleri olan DNA kompleksleri oluşturmak mümkündür. DNA normalde doğrusal bir moleküldür, ama bağlantı noktaları içeren DNA molekülleri yapılabilir. Örneğin, dört kollu bir bağlantı yapmak için birbirine uygun biçimde komplementer olan dört DNA ipliği kullanılabilir. Watson-Crick baz eşleşmesi sayesinde, ipliklerin sadece birbirine komplementer olan kısımları birbirine bağlanarak ikili DNA oluşturur. Bu dört kollu bağlantı Holliday bağlantısının hareketsiz bir tipidir.
Bağlantılar daha karmaşık molekülleri oluşturmakta kullanılabilir. Bunların en önemlisi "çifte krosover" motifidir (İngilizce literatürde buna DX kısaltmasıyla değinilir). Bunda, birbirine paralel iki DNA ikilisi vardır, bunlar iki bağlantı noktasını paylaşırlar; bu bağlantı noktalarında birer iplik bir ikiliden öbürüne geçer. Bu yapının avantajı, belli bir doğrultu ile sınırlandırılmış olmasıdır, oysa dört kollu bağlantı esnektir. Bu sayede, çifte krosover yapısı daha karmaşık kompleksler için bir yapı elemanı olmaya uygundur.[2]
  4 kollu bir bağlantı. Sol: Bir şema. Sağ: Daha gerçekçi bir model.[4] DNA'nın dört ayrı ipliği farklı renklerde gösterilmiştir. |  |
Tasarım
DNA nanoteknolojisinin ana hedeflerinden biri, belli bir hedef yapı veya işlevi sağlamak için birbirleriyle birleşecek DNA moleküllerinin dizilerini belirlemektir. Arzu edilen bir yapıyı oluşturmakta kullanılacak DNA dizilerini tasarlamak için birkaç farklı yol vardır.
Karolara dayalı yapılar
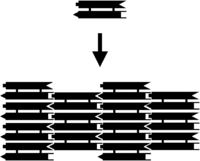
DNA nanoyapıları oluşturmanın ilk yöntemi, bunları daha küçük birimlerden inşa etmekti. Parke karolarıyla bir zeminin kaplanmasına benzeyen bu yöntemin avantajı, her bir karoyu oluşturan etkileşimler ile, tüm yapıyı oluşturan etkileşimlerin ayrı tasarlamasını mümkün kılmasıdır. Genelde periyodik örgüler yapmakta kullanılan bu yöntem, algoritmik özkurgu (İng self-assembly) gerçekleştirmekte de kullanılır, bu da DNA hesaplamasının temelini oluşturur.
Katlamalı yapılar
Karoya dayalı yaklaşıma bir alternatif olarak, uzun DNA ipliklerinin arzu edilen şekiller halinde katlanmasını sağlayarak iki boyutlu DNA yapıları oluşturmaktır. Bu katlanma, kısa "zımba" DNA ipliklerin etkisiyle sağlanır. Bu inşaat prensibine örnek olarak bir gülümseyen yüz, bir de Kuzey Amerika'nın basit bir haritası yaratılmıştır. Bu yöntem DNA origamisi olarak adlandırılmıştır.[6]
Kinetik kurgu
DNA nanoteknolojisindeki çoğu tasarım, yapının oluşumunun izlediği reaksiyon yoluna önem vermeden, hedef yapının termodinamik bir minimumda yer almasına odaklanır. Ancak, DNA özkurgusunun kinetiğini kontrolüne de önem verilebilir, kurgu sırasındaki oluşan geçici yapıların dinamikleri de programlanabilir. Bu yöntemin avantajı, yapı oluşumunun sabit sıcaklıkta gerçekleşmesi, yani tamamen termodinamik prensiplere göre yapılmış tasarımlardaki tavlama adımına gerek göstermemesidir.[7]
Dizi tasarımı
Yukarıdaki yaklaşımlardan biri ile hedef molekülün ikincil yapısı tasarladıktan sonra, arzu edilen yapıyı oluşturacak bir nükleotit dizisinin tasarlanması gerekir. Nükleik asit tasarımı, arzu edilen bir biçimde (örneğin bakınız RNA yapısı) birleşecek nükleik asit baz dizilerinin üretimidir. Nükleik asit tasarımı DNA nanoteknoloji sahasında merkezi bir yeri vardır.
Nükleik asit tasarımının amacı protein tasarımınkine benzer: ikisinde de, arzu edilen yapının oluşmasına eğilimli olan ve arzu edilmeyen yapıları oluşmasına eğilimli olmayan monomerler dizisi tasarlanır. Hesaplama yapmak açısından nükleik asit tasarımı daha basit bir problemdir, çünkü Watson-Crick baz eşleşme kurallarının sadeliği, basit buluşsal yöntemleri mümkün kılar, bunlardan deneysel anlamda sağlam tasarımlar elde edilir. Ancak, nükleik asit yapıları, proteinlere kıyasla işlevleri bakımından daha sınırlıdır. [8][9]
Hedef yapılar
DNA'dan oluşan pek çok yapı sentezlenmiş ve karakterize edilmiştir.
İki boyutlu örgüler

çifte krosover (DX) molekülleri yapışkan uçlara sahip olurlarsa bunlar birbiriyle birleşerek iki boyutlu periyodik bir örgü (latis) oluşturabilirler. Her bir DX molekülünün dört ucu vardır, bunlar iki çifte sarmal bölgenin uçlarında yer alırlar. Bu uçlar yapışkan olursa bunların arzu edilen belli biçimlerde birleşmesi sağlanabilir. Birden çok DX tipi kullanılarak bunların diziler veya başka tesselasyon biçimleri olarak düzenlenebilir. Meydana gelen yassı yapraklar aslında iki boyutlu DNA kristalleridirler.[11][12]
İki boyutlu dizilimler başka motifler kullanılarak da yapılmıştır. Bunların arasında Holliday bağlantı eşkenar dörtgen dizilimi ve, üçgen veya altıgen şekilli çeşitli çifte krosover-temelli dizilimler sayılabilir.[13][14][15]

Ayrık üç boyutlu yapılar
Küp veya tetrahedron gibi çokyüzlülere benzer bir takım DNA moleküllü yapılar oluşturulmuştur. Bir diğer deyişle, DNA ikilileri bir çokyüzlünün kenarlarını, DNA bağlantıları da köşelerini oluşturur. DNA çokyüzlüsünün en erken gösteriminde, çok sayıda ligasyon ve katı hâl sentez adımı ile bir kafes çokyüzlü yaratılmıştır.[17] Diğer örnekler, doğru şekilde katlanacak şekilde tasarlanmış uzun bir iplikten yapılmış köşeleri kesik bir oktahedron ve dört DNA ipliğinden tek bir adımda meydana gelebilen bir tetrahedrondur.[16][18]
Katı yüzlü DNA yapıları da inşa edilmiştir, DNA origami yöntemi ile. Bu yapılar bir uyarı ile açılıp içlerinde taşıdıkları kargoyu boşaltacak şekilde programlanabilirler, böylece bunların programlanabilir moleküler kafes uygulamaları olabilir[19][20]
DNA nanotüpleri
Yassı yapraklara ek olarak, çifte krosover dizilimlerinin, 4-20 nm çapında içi boş tüpler oluşturması da sağlanabilmiştir. Bu DNA nanotüpleri karbon tüplerine benzer büyüklük ve boyuttadırlar ama karbon nanotüpleri daha sağlam ve daha iletkendir, buna karşın DNA nanotüplerini daha kolay modifiye edilebilir ve başka yapılara bağlanabilir.[21]
Genişletilmiş üç boyutlu yapılar
DNA'dan üç boyutlu örgüler yapmak DNA nanoteknolojisinin ilk amaçlarından biriyid ama tahmin edilenden daha zor oldu. Üç boyutlu örgülerin inşasında başarı nihayet 2009'da, tensegrite kavramina dayalı bir motif ile mümkün oldu. Tensegrite, gerilim ile sıkışma güçleri arasındaki dengedir.[22]
Uygulamalar
DNA nanoteknolojisi tasarlanmış işlevlere ve yapılara sahip moleküller yaratmayı amaçlar. Çeşitli işlevsel sınıflar gösterilmiştir.
Nanomimari
DNA dizilimlerini bir kalıp gibi kullanıp başka işlevsel moleküllerin kurgusunu sağlamak fikri ilk defa Nadrian Seeman tarafından 1987'de önerilmiş,[23] ama ancak 2006'da bu planların pratik uygulaması gösterilebildi. DX temelli bir karoya bir altın nanotaneciği bağlanmış ve DNA yapılarının kendi kendini oluşturmasının (öz kurgusunun) sonucunda DNA'lara bağlı olan nanotaneciklerin de birleştiği gösterilmişir. Bir diğer uygulamada, DX dizilimi üzerinde Dervan poliamidleri kullanılarak, DNA diziliminde yer alan belli çeşit karolar üzerinde streptavidin moleküllerinin yerleştirilmesi sağlanabilmiştir.[24][25]
2006'da Dwyer ve LaBean 4x4'lük bir DX dizilimi üzerinde, streptavidin kullanarak, "D" "N" and "A" harfleri oluşturulmuştu.[26] 2007'de, hiyerarşik bir kurgu yöntemi ile bu yaklaşım daha büyük dizilimlere de uygulanmıştır (8X8 ve 8.96 Mega Dalton kütleli).[27]
DNA nanoteknolojisi ile moleküler elektronik cihazlar kurma girişimleri de yapılmıştır. Bu amaçla, DNA kullanılarak tek duvarlı karbon nanotüplü alan etkili transistör oluşturulmuştur.[28]
Algoritmik özkurgu


DNA nanoteknolojisi, ilgili bir saha olan DNA hesaplamasına da uygulanmıştır. DX karolarının yapışkan uçları uygun şekilde seçilirse bunlar Wang karoları gibi davranıp hesaplama yapabilir. Bir DX diziliminin bir XOR mantık işlemini kodladığı gösterilmiştir. B sayede, bir DNA dizilimi bir hücresel otomat (cellular automaton) oluşturmakta, bu da, Sierpinski üçgeni olarak adlandırılan bir fraktal üretmektedir. Böylece bir DNA dizilimine hesaplama işlevinin de dahil edilebileceği gösterilmiştir.
DNA hesaplaması ile DNA nanoteknolojisi örtüşen sahalar olmakla beraber ayrıktırlar. DNA nanoteknolojisi, Watson-Crick baz eşleşmesinin spesifisitesini kullanarak DNA'dan yeni, değişik yapılar meydana getirir. Bu yapılar DNA hesaplaması için kullanılabilir ama böyle bir şart yoktur. Buna ek olarak, DNA nanoteknolojisi ile mümkün kılınmış moleküller olmadan da DNA hesaplaması yapmak mümkündür.[29]
DNA nanomekanik cihazlar
Belli bir dürtü sonucu şeklini değiştirebilen DNA kompleksleri imal edilmiştir. Bunlar nanorobotik uygulamaları için tasarlanmıştır. Bu tür cihazların ilk yapılanlarından olan "moleküler cımbız", bir kontrol ipliğinin etkisiyle açık bir hâlden kapalı bir hâle geçebilmektedir.[30]
Burulma hareketi yapabilen DNA kompleksleri de imal edilmiştir. Bunlar B-DNA ile Z-DNA biçimleri arasındaki geçiş yaparak çözeltideki değişen şartlara cevap vermektedir.[31] Bir diğeri, bir kontrol ipliğinden yararlanarak, paranemik krosover konformasyonundan çifte bağlantı konformasyonuna geçiş yapmaktadır.[32]
Malzeme ve yöntemler
Arzu edilen diziye sahip DNA molekülleri oligonükleotit sentezi ile kolayca elde edilebilir. Bu işlem genelde bir DNA sentezleme makinası kullanılarak otomatikleştirilmiştir ve çoğu şirket aracılığıyla siparişli DNA elde edilebilir.
Hedef yapıyı olşturacak DNA ipliklerinin dizileri bilgisayarla tasarlanır. Moleküler modelleme ve termodinamik modelleme DNA dizilerini optimize etmek için bazen kullanılır.
DNA nanoteknolojisinde kullanılan DNA molekülleri genelde jel elektroforezi ile karakterize edilir. Böylece DNA moleküllerinin büyüklüğü ve şekli hakkında bilgi elde edilir ve bu moleküllerin doğru oluştukları kontrol edilir. Bu moleküllerin yaplarını karakterize etmek için flüoresan işaretleme ve Flüresans rezonans enerji transferi
DNA yapılarını görüntülemek için atomik güç mikroskopisi kullanılır, bu yöntem ile düz yüzeyler üzerindeki yapıların resimleri çekilebilir. Bu araç iki boyutlu kristaller için uygundur ama ayrık üç boyutlu yapılar için pek yararlı değildir. Bu tür yapılar için kriyo-elektron mikroskopi popülerleşen bir yöntemdir. Büyük üç boyutlu yapıların analizi X-ışını kristalografisi ile yapılır. DNA özkurgusunun kinetiğini araştırmak için ikili polarizasyon enteroferometrisi ve QCMD kullanılabilir.
Ayrıca bakınız
- DNA yapısı
- DNA'nın moleküler modelleri
Dış başlantılar
- Üç boyutlu DNA tasarımı, modelleme ve/veya benzetimi için yazılımlar:
- Ascalaph Designer
- caDNAno 7 Ağustos 2011 tarihinde Wayback Machine sitesinde arşivlendi.
- GIDEON 6 Ağustos 2020 tarihinde Wayback Machine sitesinde arşivlendi.
- NanoEngineer-1 13 Eylül 2019 tarihinde Wayback Machine sitesinde arşivlendi.
- Uluslararası Nano-ölçekli Bilim, Hesaplama ve Mühendislik Derneği [1] 29 Temmuz 2012 tarihinde Wayback Machine sitesinde arşivlendi.
Kaynakça
- ^ a b c Pelesko, John A. (2007). Self-assembly: the science of things that put themselves together. New York: Chapman & Hall/CRC. ss. 201, 242, 259. ISBN 978 1 58488 687 7.
- ^ a b c d Seeman, Nadrian C. (Haziran 2004). "Nanotechnology and the double helix". Scientific American. 290 (6). ss. 64-75. doi:10.1038/scientificamerican0604-64. PMID 15195395. 10 Kasım 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2010.
- ^ Problemin sunumu için Nadrian Seeman'ın Web sitesindeki, kristalizasyon protokolüne 30 Kasım 2012 tarihinde Wayback Machine sitesinde arşivlendi. bakınız, önerilen bir çözüm için de gene Nadrian Seeman's sitesinde, yönledirilmiş konuklar içeren DNA kafesleri 1 Kasım 2016 tarihinde Wayback Machine sitesinde arşivlendi. sayfasına bakınız.
- ^ PDB 1M6G 10 Ocak 2010 tarihinde Wayback Machine sitesinde arşivlendi. koordinatlarından yaratılmıştır
- ^ a b Mao, Chengde (Aralık 2004). "The Emergence of Complexity: Lessons from DNA". PLoS Biology. 2 (12). ss. 2036-2038. doi:10.1371/journal.pbio.0020431. ISSN 1544-9173.
- ^ Rothemund, Paul W. K. (2006). "Folding DNA to create nanoscale shapes and patterns". Nature. Cilt 440. ss. 297-302. doi:10.1038/nature04586. ISSN 0028-0836.
- ^ Yin, Peng; Choi, Harry M. T.; Calvert, Colby R.; Pierce, Niles A. (2008). "Programming biomolecular self-assembly pathways". Nature. 451 (7176). s. 318. doi:10.1038/nature06451. PMID 18202654.
- ^ Dirks, Robert M. (2004). "Paradigms for computational nucleic acid design". Nucleic Acids Research. 32 (4). ss. 1392-1403. doi:10.1093/nar/gkh291. PMC 390280 $2. PMID 14990744.
- ^ Dirks, Robert M.; Bois, Justin S.; Schaeffer, Joseph M.; Winfree, Erik; Pierce, Niles A. (2007). "Thermodynamic Analysis of Interacting Nucleic Acid Strands". SIAM Review. Cilt 49. s. 65. doi:10.1137/060651100.
- ^ Strong, Michael (2004). "Protein Nanomachines". PLoS Biology. 2 (3). ss. e73. doi:10.1371/journal.pbio.0020073. PMC 368168 $2. PMID 15024422.
- ^ Winfree, Erik (6 Ağustos 1998). "Design and self-assembly of two-dimensional DNA crystals". Nature. Cilt 394. ss. 529-544. doi:10.1038/28998. ISSN 0028-0836.
- ^ Liu, Furong (10 Şubat 1999). "Modifying the Surface Features of Two-Dimensional DNA Crystals". Journal of the American Chemical Society. 121 (5). ss. 917-922. doi:10.1021/ja982824a. ISSN 0002-7863.
- ^ Mao, Chengde (16 Haziran 1999). "Designed Two-Dimensional DNA Holliday Junction Arrays Visualized by Atomic Force Microscopy". Journal of the American Chemical Society. 121 (23). ss. 5437-5443. doi:10.1021/ja9900398. ISSN 0002-7863.
- ^ Constantinou, Pamela E. (2006). "Double cohesion in structural DNA nanotechnology". Organic and Biomolecular Chemistry. Cilt 4. ss. 3414-3419. doi:10.1039/b605212f.
- ^ Mathieu, Frederick (Nisan 2005). "Six-Helix Bundles Designed from DNA". Nano Letters. 5 (4). ss. 661-665. doi:10.1021/nl050084f. ISSN 1530-6984.
- ^ a b Goodman, R.P. (9 Aralık 2005). "Rapid chiral assembly of rigid DNA building blocks for molecular nanofabrication". Science. 310 (5754). ss. 1661-1665. doi:10.1126/science.1120367. ISSN 0036-8075.
- ^ Zhang, Yuwen (1994). "Construction of a DNA-truncated octahedron". Journal of the American Chemical Society. 116 (5). ss. 1661-1669. doi:10.1021/ja00084a006. ISSN 0002-7863.
- ^ Shih, William M. (12 Şubat 2004). "A 1.7-kilobase single-stranded DNA that folds into a nanoscale octahedron". Nature. Cilt 427. ss. 618-621. doi:10.1038/nature02307. ISSN 0028-0836.
- ^ Andersen, Ebbe S.; Dong, Mingdong; Nielsen, Morten M.; Jahn, Kasper; Subramani, Ramesh; Mamdouh, Wael; Golas, Monika M.; Sander, Bjoern; Stark, Holger (2009). "Self-assembly of a nanoscale DNA box with a controllable lid". Nature. 459 (7243). s. 73. doi:10.1038/nature07971. PMID 19424153.
- ^ Ke, Yonggang; Sharma, Jaswinder; Liu, Minghui; Jahn, Kasper; Liu, Yan; Yan, Hao (2009). "Scaffolded DNA Origami of a DNA Tetrahedron Molecular Container". Nano Letters. 9 (6). s. 2445. doi:10.1021/nl901165f. PMID 19419184.
- ^ Rothemund, Paul W. K. (22 Aralık 2004). "Design and Characterization of Programmable DNA Nanotubes". Journal of the American Chemical Society. 126 (50). ss. 16344-16352. doi:10.1021/ja044319l. ISSN 0002-7863.
- ^ Zheng, Jianping; Birktoft, Jens J.; Chen, Yi; Wang, Tong; Sha, Ruojie; Constantinou, Pamela E.; Ginell, Stephan L.; Mao, Chengde; Seeman, Nadrian C. (2009). "From molecular to macroscopic via the rational design of a self-assembled 3D DNA crystal". Nature. 461 (7260). s. 74. doi:10.1038/nature08274. PMC 2764300 $2. PMID 19727196.
- ^ Robinson, Bruche H. (Ağustos 1987). "The Design of a Biochip: A Self-Assembling Molecular-Scale Memory Device". Protein Engineering. 1 (4). ss. 295-300. doi:10.1093/protein/1.4.295. ISSN 0269-2139. PMID 3508280. 25 Mayıs 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2010.
- ^ Zheng, Jiwen (2006). "2D Nanoparticle Arrays Show the Organizational Power of Robust DNA Motifs". Nano Letters. Cilt 6. ss. 1502-1504. doi:10.1021/nl060994c. ISSN 1530-6984.
- ^ Cohen, Justin D. (2007). "Addressing Single Molecules on DNA Nanostructures". Angewandte Chemie. 46 (42). ss. 7956-7959. doi:10.1002/anie.200702767. ISSN 0570-0833.
- ^ Park, Sung Ha (Ekim 2006). "Finite-Size, Fully Addressable DNA Tile Lattices Formed by Hierarchical Assembly Procedures". Angewandte Chemie. 118 (40). ss. 749-753. doi:10.1002/ange.200690141. ISSN 1521-3757. 25 Mayıs 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2010.
- ^ Pistol, Constantin (Mart 2007). "Scalable, Low-cost, Hierarchical Assembly of Programmable DNA Nanostructures". Nanotechnology. 18 (12). ss. 125305-9. doi:10.1088/0957-4484/18/12/125305. ISSN 0957-4484.
- ^ Keren, K. (Kasım 2003). "DNA-Templated Carbon Nanotube Field-Effect Transistor". Science. 302 (6549). ss. 1380-1382. doi:10.1126/science.1091022. ISSN 1095-9203. 17 Eylül 2012 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Haziran 2010.
- ^ a b Rothemund, Paul W. K. (Aralık 2004). "Algorithmic Self-Assembly of DNA Sierpinski Triangles". PLoS Biology. 2 (12). ss. 2041-2053. doi:10.1371/journal.pbio.0020424. ISSN 1544-9173.
- ^ Yurke, Bernard (10 Ağustos 2000). "A DNA-fuelled molecular machine made of DNA". Nature. Cilt 406. ss. 605-609. doi:10.1038/35020524. ISSN 0028-0836.
- ^ Mao, Chengde (14 Ocak 1999). "A DNA Nanomechanical Device Based on the B-Z Transition". Nature. Cilt 397. ss. 144-146. doi:10.1038/16437. ISSN 0028-0836.
- ^ Yan, Hao (3 Ocak 2002). "A robust DNA mechanical device controlled by hybridization topology". Nature. Cilt 415. ss. 62-65. doi:10.1038/415062a. ISSN 0028-0836.