DNA ligaz

Moleküler biyolojide DNA ligaz iki DNA molekülünü uç uca birleştiren özel bir ligaz (EC 6.5.1.1) tipidir. DNA ligaz DNA tamiri, DNA ikileşmesinde rol oynar. Ayrıca, ökaryotlarda mayoz bölünmedeki krosoverde ve memelilerde, bağışıklık sisteminin çeşitliliğini sağlayan rekombinasyon süreçlerinde rol oynarlar. DNA ligaz enzimi moleküler biyoloji laboratuvarlarında rekombinant DNA uygulamalarında kullanılır.
Ligaz mekanizması
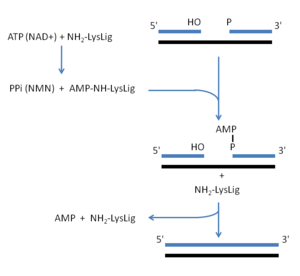
DNA ligazın görevi bir DNA molekülünün 3' hidroksil ucu ile bir diğerinin 5' fosfat ucu arasında bir fosfodiester bağ oluşturmaktır. Katalizledikleri reaksiyon için yüksek enerjili bir kofaktör olarak ATP veya NAD kullanırlar. Ökaryotik DNA ligazlar ATP ile çalışır, bakteri, arke ve virüslerdeki DNA ligazların bazıları ATP bazıları NAD kullanır. Bu kofaktörlerin fosfat bağının kesilmesi ile reaksiyon için gerekli enerji elde edilir. Arta kalan AMP molekülü enzimde bulunan bir lizin kalıntısı tarafından bağlanır. Birleştirilecek DNA molekülünün 5' ucundaki fosfat, lizine bağlı AMP'deki 5' fosfata saldırarak, AMP'yi DNA'ya bağlayan bir pirofosfat bağı oluşturur. Nihayet, kopuk DNA zincirinin 3'-OH grubu, 5' uçtaki pirofosfat grubuna saldırarak AMP'nin serbest bırakılmasına ve DNA zincir uçlarının birleşmesine neden olur.[1]
DNA ligazlar DNA'yı çevreleyerek ona şekil verirler ve birleşmesi gereken zincir uçlarını karşı karşıya getirirler.
Tüm DNA ligazlarda DNA'ya bağlanan bir bölge, bir de nükleotidil transferaz bölgesi vardır. Nükleotidil transferaz bölgesi AMP'yi DNA'ya aktarır. Bu iki bölgeye ek olarak, bazı DNA ligazlarda yardımcı proteinlerle etkileşmeyi sağlayan ek bölgeler bulunur. Diğer DNA ligazlardan farklı olarak DNA ligaz III'te DNA bağlanan iki ayrı bölge vardır, bu sayede küt uçlu iki DNA uçuca gelebilir.[2]
DNA ligazlarda görülen, yukarıda belirtilen üç adımlı mekanizma RNA ligazlarda da mevcuttur. Mesajcı RNA'ya 5' başlık takan enzimler ise bu üç adımın ilk ikisini sadece uygular ve DNA yerine RNA, ATP yerine GTP kullanır. Bu enzimlerin aktif bölgelerinin yapısı da benzerdir. Katalizledikleri reaksiyon adımlarından biri, bir nükleotitin bir nükleik aside eklenmesi olduğu için bu enzim aileleri toplu olarak nükleotidiltransferaz enzim üst ailesini oluştururlar.[2]
DNA ligaz tipleri
NAD ve ATP kullanan DNA ligazların biyokimyasal mekanizmaları aynı olmasına karşın amino asit dizileri bakımından birbirlerinden çok farklıdır ve iki ayrı grup oluştururlar.
Bakteriyel DNA ligazlar
E. coli DNA ligazı DNA ikileşmesi sırasında oluşan Okazaki parçalarının birleştirilmesi, ayrıca DNA tamiri için gereklidir. Bakteriyofaj T4 ve T7'de bulunan DNA ligazlar da aynı göreve sahiptir ama konak bakteride bulunan DNA ligaz, faj DNA'sının ikileşmesinde işlev görebildiği için onların varlıkları faj çoğalması için zorunlu değildir.[3]
Ökaryotlarda DNA ligazları
İnsanda LIG1, LIG3 ve LIG4 genleri tarafından kodlanan üç DNA ligaz geni vardır. LIG1 ve LIG4 tüm ökaryotlarda bulunmakta, LIG3 ise sadece omurgalı hayvanlarda bulunmaktadır.[2]
- DNA ligaz I, DNA ikileşmesi sırasında Okazaki parçalarını birleştirir. Ayrıca morötesi kaynaklı DNA hasarının tamirinde görev alır ama bu sadece çoğalmakta olan hücrelerde olur. Durağan hücrelerde DNA ligaz III bu işi yapar.[4]
- DNA ligaz II, bölünmeyen hücrelerde görülür, DNA ligaz III'ün alternatif uçbirleştirme ürünüdür.
- DNA ligaz III, DNA tamir proteini XRCC1 ile kompleksleşerek baz eksizyon tamirinin son aşamasında fosfodiester bağını onarır. Ayrıca, mayoz bölümedeki rekombinasyonda rol oynar.[5] DNA ligaz III homologları tüm omurgalı hayvanlarda bulunur.
- DNA ligaz IV, XRCC4 adlı yardımcı protein ile kompleksleşir. Çift zincirli DNA tamirinin son adımını katalizler. Ayrıca, bağışıklık sistemi gelişiminde immünoglobin ve T-hücre reseptöründe çeşitlilik yaratan V(D)J rekombinasyonu için gereklidir. Bu süreçte DNA ligaz IV, non-homolog DNA uç birleştirme reaksiyonunu katalizler.
Sağlık
DNA ligaz enzimlerinde yokluğu veya doğru çalışmaması sonucu genom yapısında bozukluklar meydana gelir. DNA ligaz I mutasyonu taşıyan hastalarda DNA ikileşmesi ve tamiri iyi çalışmamasından dolayı immün bozukluklar, güneş ışığına duyarlılık ve büyüme gecikmesi görülmüştür. Memeli hayvanlarda DNA ligaz I veya IV eksikliği kanser olasılığını artırdığı gösterilmiştir. DNA ligaz IV yokluğu sendromunda V(D)J rekombinasyonu bozukluğundan kaynaklanan immün yetmezlik görülür. Fare modellerinde ve çoğu insan hastada DNA ligaz IV yokluğu kansere yol açmaktadır.[2]
Uygulama

DNA ligazlar, laboratuvarda rekombinant DNA dizileri oluşturmak için vazgeçilmez birer araç haline gelmişlerdir. Örneğin restriksiyon enzimleri ile elde edilen DNA parçalarının bir plazmit içine yerleştirilebilmesi için DNA ligazlar kullanılır. Uçları birleştirilen DNA moleküllerinin uçları "yapışkan" (sağdaki resimdeki gibi) veya küt olabilir.
Başarılı bir ligasyon (birleştirme) reaksiyonu için en iyi sıcaklığın bulunması gereklidir. Çoğu reaksiyon bakteriofaj T4'ten elde edilen T4 DNA ligazı ile katalizlenir. Ancak ligasyonun başarılı olması için enzim etkinliği ile DNA uçlarının ergimesi arasında bir denge bulunması gerekir. Eğer reaksiyon sıcaklığı ergime sıcaklığı (buna tavlama sıcaklığı da denir) Tm'den yüksek olursa, yapışkan uçlar arasında baz eşleşmesi olmaz. Yapışkan uçların uzantısı ne kadar kısa olursa Tm de o kadar düşük olur. Bu yüzden kısa uzantılı yapışkan uçların ligasyonu düşük sıcaklıkta (4-8 °C) uzun süre içinde (saatlerce, genelde bir gece boyunca) gerçekleştirilir.
5' ucunda fosfat olmayan DNA'ların ligaz tarafından birleştirilebilmesi için 5' uçlarının fosforile edilmesi gerekir.
Yapışkan uçsuz DNA'ların da ligasyonu mümkündür ama reaksiyon çok daha yavaştır. Reaksiyon ortamında polietilen glikol ve 150-200 mM NaCl bulunması DNA moleküllerinin bitişmesini kolaylaştırır.
Kaynakça
- ^ Lehman, I. R. DNA ligase: structure, mechanism, and function. Science 186, 790−797 (1974)
- ^ a b c d Tom Ellenberger, T., Tomkinson A.E. (2008) Eukaryotic DNA Ligases: Structural and Functional Insights. Ann. Review of Biochemistry 77: 313-338
- ^ C.W. Fuller, B.B. Beauchamp, M.J. Engler, R.L. Lechner, S.W. Matson, S. Tabor, J.H. White, C.C Richardson. Mechanisms for the Initiation of Bacteriophage T7 DNA Replication. Cold Spring Harb Symp Quant Biol 1983. 47: 669-679
- ^ Moser, J.; Kool, H.; Giakzidis, I.; Caldecott, K.; Mullenders, L. H. F.; Fousteri, M. I. : Sealing of chromosomal DNA nicks during nucleotide excision repair requires XRCC1 and DNA ligase III-alpha in a cell-cycle-specific manner. Molec. Cell 27: 311-323, 2007. PubMed ID : 17643379
- ^ Chen, J.; Tomkinson, A. E.; Ramos, W.; Mackey, Z. B.; Danehower, S.; Walter, C. A.; Schultz, R. A.; Besterman, J. M.; Husain, I. : Mammalian DNA ligase III: molecular cloning, chromosomal localization, and expression in spermatocytes undergoing meiotic recombination. Molec. Cell. Biol. 15: 5412-5422, 1995. PubMed ID : 7565692
Dış bağlantılar
- DNA Ligase: PDB ayın molekülü (İngilizce)
- Ligaz reaksiyonunun kimyasal adımları2 Nisan 2015 tarihinde Wayback Machine sitesinde arşivlendi. (İngilizce)