Crenarchaeota
| Crenarchaeota | |||||
|---|---|---|---|---|---|
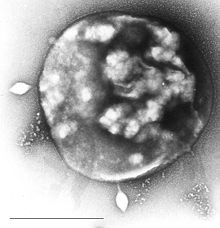 Belirli bir virüs STSV-1 ile enfekte Archaea Sulfolobus | |||||
| Korunma durumu | |||||
| Değerlendirilmedi (IUCN 3.1) | |||||
| Biyolojik sınıflandırma | |||||
| |||||
| Sinonimler | |||||
| |||||
Crenarchaeota, Archaea âlemine bağlı ökaryotlarla ilişkili bir algae şubesidir.[2][3][4][5] Başlangıçta, Crenarchaeota'nın kükürt bağımlı ekstremofiller olduğu düşünülüyordu, ancak son çalışmalar, organizmaların deniz ortamında en bol bulunan arke olabileceğini gösteren karakteristik Crenarchaeota çevresel rRNA'sını tanımladı.[6] Başlangıçta, rRNA dizilerine dayalı olarak diğer arkelerden ayrılmışlardı; bazı crenarchaea'ların histonlara sahip olduğu tespit edilmesine rağmen, histon eksikliği gibi diğer fizyolojik özellikler bu bölünmeyi desteklemiştir.[7] Yakın zamana kadar kültürlenmiş tüm Crenarchaealar, bazıları 113 °C'ye kadar büyüme yeteneğine sahip olan termofilik veya hipertermofilik organizmaların.[8] Bu organizmalar gram negatif boyanır ve morfolojik olarak çeşitlidir; çubuk, kok, ipliksi ve tuhaf şekilli hücrelere sahiptir.[9]
Taksonomi
Crenarchaeota şubesine bağlı sınıflar (2022):
Sulfolobus
Crenarcheota'nın en iyi karakterize edilmiş üyelerinden biri Sulfolobus solfataricus'tur. Bu organizma orijinal olarak İtalya'daki jeotermal olarak ısıtılan sülfürik kaynaklardan izole edilmiştir ve 80 °C'de ve 2-4 pH'da büyür.[10] Termofil ve arke araştırmalarında bir öncü olan Wolfram Zillig tarafından ilk karakterizasyonundan bu yana, aynı cinse ait benzer türler dünya çapında bulunmuştur. Kültürlü termofillerin büyük çoğunluğunun aksine, Sulfolobus aerobik ve kemoorganotrofik olarak büyür (enerjisini şekerler gibi organik kaynaklardan alır). Bu faktörler, laboratuvar koşullarında anaerobik organizmalardan çok daha kolay bir büyümeye izin verir ve Sulfolobus'un hipertermofiller ve bunların içinde çoğalan çok çeşitli virüslerin incelenmesi için model bir organizma haline gelmesine yol açmıştır.
Deniz türleri
1992'den başlayarak, deniz ortamlarında Crenarchaea'ya ait gen dizilerini bildiren veriler yayınlandı.[11][12] O zamandan beri, açık okyanustan alınan Crenarchaea zarlarından bol miktarda lipidlerin analizi, bu “düşük sıcaklıklı Crenarchaea” konsantrasyonunu belirlemek için kullanılmıştır (Bkz. TEX-86). İmza lipitlerinin bu ölçümlerine dayanarak, Crenarchaea'nın çok bol olduğu ve karbonun sabitlenmesine ana katkıda bulunanlardan biri olduğu düşünülmektedir.[] Crenarchaea'dan gelen DNA dizileri toprak ve tatlı su ortamlarında da bulunmuştur, bu da bu filumun çoğu ortamda her yerde bulunduğunu düşündürmektedir.[13]
2005 yılında, ilk kültürlü "düşük sıcaklıklı Crenarchaea”nın kanıtı yayınlandı. Nitrosopumilus maritimus adlı, bir deniz akvaryumu tankından izole edilen ve 28 yaşında yetiştirilen amonyak oksitleyici bir organizmadır. °C.[14]

Eosit hipotezi
1980'lerde James Lake tarafından önerilen eosit hipotezi, ökaryotların prokaryotik eositler içinde ortaya çıktığını öne sürer.
Crenarchaea ve ökaryotlar arasında yakın bir ilişkiyi destekleyen olası bir kanıt, Crenarchea'da RNA polimeraz alt birimi Rbp-8'in bir homologunun varlığıdır, ancak bu Euryarchaeota'da bulunmaz.[16]
Ayrıca bakınız
Dış bağlantılar
 Wikimedia Commons'ta Crenarchaeota ile ilgili çoklu ortam belgeleri bulunur.
Wikimedia Commons'ta Crenarchaeota ile ilgili çoklu ortam belgeleri bulunur. Vikitür'de Crenarchaeota ile ilgili ayrıntılı taksonomik bilgiler bulunur.
Vikitür'de Crenarchaeota ile ilgili ayrıntılı taksonomik bilgiler bulunur.
Kaynakça
- ^ "Crenarchaeota". itis.gov.
- ^ "ITIS". 30 Ekim 2013 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Temmuz 2022.
- ^ See the NCBI webpage on Crenarchaeota 29 Ocak 2019 tarihinde Wayback Machine sitesinde arşivlendi.
- ^ C.Michael Hogan. 2010. Archaea. eds. E.Monosson & C.Cleveland, Encyclopedia of Earth. National Council for Science and the Environment, Washington DC. 11 Mayıs 2011 tarihinde Wayback Machine sitesinde arşivlendi.
- ^ Data extracted from the "NCBI taxonomy resources". National Center for Biotechnology Information. 12 Ekim 2012 tarihinde kaynağından arşivlendi. Erişim tarihi: 19 Mart 2007.
- ^ Brock Biology of Microorganisms. 11th. Prentice Hall. 2005. ISBN 978-0-13-144329-7.
- ^ "Histones in Crenarchaea". Journal of Bacteriology. 187 (15): 5482-5485. 2005. doi:10.1128/JB.187.15.5482-5485.2005. PMC 1196040 $2. PMID 16030242.
- ^ "Pyrolobus fumarii, gen. and sp. nov., represents a novel group of archaea, extending the upper temperature limit for life to 113 °C". Extremophiles. 1 (1): 14-21. 1997. doi:10.1007/s007920050010. PMID 9680332.
- ^ Bergey's Manual of Systematic Bacteriology Volume 1: The Archaea and the Deeply Branching and Phototrophic Bacteria. 2nd. Springer. 2001. ISBN 978-0-387-98771-2.
- ^ "The Sulfolobus-"Caldariellard" group: Taxonomy on the basis of the structure of DNA-dependent RNA polymerases". Arch. Microbiol. 125 (3): 259-269. 1980. doi:10.1007/BF00446886.
- ^ "Novel major archaebacterial group from marine plankton". Nature. 356 (6365): 148-9. 1992. doi:10.1038/356148a0. PMID 1545865.
- ^ DeLong EF (1992). "Archaea in coastal marine environments". Proc Natl Acad Sci USA. 89 (12): 5685-9. doi:10.1073/pnas.89.12.5685. PMC 49357 $2. PMID 1608980.
- ^ "Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences". Proc Natl Acad Sci USA. 93 (17): 9188-93. 1996. doi:10.1073/pnas.93.17.9188. PMC 38617 $2. PMID 8799176.
- ^ "Isolation of an autotrophic ammonia-oxidizing marine archaeon". Nature. 437 (7058): 543-6. 2005. doi:10.1038/nature03911. PMID 16177789.
- ^ Cox, C. J. (2008). "The archaebacterial origin of eukaryotes". Proc Natl Acad Sci USA. 105 (51): 20356-61. doi:10.1073/pnas.0810647105. PMC 2629343 $2. PMID 19073919.
- ^ Kwapisz (2008). "Early evolution of eukaryotic DNA-dependent RNA polymerases". Trends Genet. 24 (5): 211-5. doi:10.1016/j.tig.2008.02.002. PMID 18384908.