Biyomoleküler yapı
Biyomoleküler yapı biyomoleküllerin (başlıca protein, DNA ve RNA'nın) yapısıdır. Bu moleküllerin yapısı genelde birincil, ikincil, üçüncül ve dördüncül yapı olarak ayrılır. Bu yapının iskeleti, molekül içinde birbirine hidrojen bağları ile bağlanmış ikincil yapı elemanları tarafından oluşturulur. Bunun sonucunda protein ve nükleik asit yapı bölgeleri (İngilizce domain) oluşur.
Birincil, ikincil, üçüncül ve dördüncül yapı terimleri ilk defa Kaj Ulrik Linderstrøm-Lang tarafından Stanford Üniversitesi'ndeki 1951 Lane Tıp Konferanslarında kullanılmıştır.
Birincil yapı
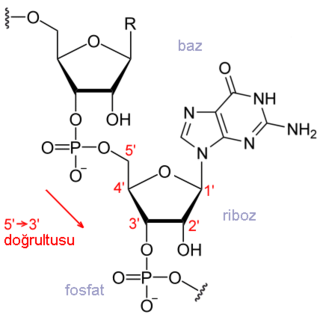
Biyokimyada bir biyomolekülün birincil yapısı, onun atomik bileşiminin ve bu atomları birbirine bağlayan kimyasal bağların (stereokimyaları da dahil olmak üzere) tam olarak belirtilmesidir. Tipik, dallanmamış, çapraz bağlanmamış bir biyopolimerde (DNA, RNA veya tipik bir protein gibi), birincil yapı, onun monomerlerinin dizisine (yani nükleotit veya peptit dizisine) karşılık gelir.
Birincil yapı bazen hatalı olarak birincil dizi olarak ifade edilir, ama böyle bir terim yoktur, keza paralel kavramlar olarak ikinci veya üçüncül dizi de yoktur. Konvansiyona göre, bir proteinin birincil yapısı onun amino (N) ucundan karboksi (C) ucunda doğru belirtilir, bir DNA veya RNA molekülünün birinci yapısı ise 5' ucundan 3' ucunda doğru beliritilir.
Bir nükleik asidin birincil yapısı, molekülü oluşturan nükleotitlerin dizisine karşılık gelir. Birincil yapıda bulunan dizi motiflerinin işlevsel önemi vardır. Bazı dizi motif örnekleri: snoRNA'ların C/D[1] ve H/ACA kutuları[2] splisozomal RNA'larda bulunan Sm bağlanma yeri, Shine-Dalgarno dizisi,[3] Kozak konsensus dizisi[4] ve RNA polymerase III sonlandırıcısıdır.[5]
İkincil yapı
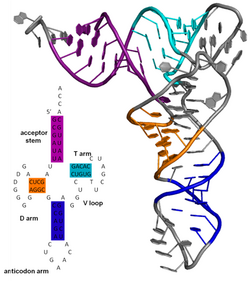
Biyokimya ve yapısal biyolojide, ikincil yapı, protein veya nükleik asit (DNA/RNA) gibi biyopolimerlerin yerel parçalarının üç boyutlu şekilleridir. Ancak üç boyutlu uzaydaki spesifik atom konumlarını belirtmez, bunlar üçüncül yapıya ait sayılır. İkincil yapı biyopolimerdeki hidrojen bağları tarafından belirlenir. Proteinlerde ikincil yapı omurgadaki amit ve karboksi grupları arasındaki hidrojen bağları tarafından belirlenir (Yan zincir-ana zincir ve yan zincir-yan zincir ilişkileri önemsizdir), nükleik asitlerde ise ikincil yapı azotlu bazlar arasındaki hidrojen bağları tarafından belirlenir.
Proteinlerde hidrojen bağlanması diğer yapısal özellikler ile ilişkilidir, bu yüzden ikincil yapının daha az resmî bir tanımlamasına yol açmıştır. Örneğin, protein sarmallarındaki amino asit kalıntıları genelde Ramachandran grafiğinin belli bir bölgesindeki omurga dihedral açılarına sahiptir; dolayısıyla bu dihedral açılara sahip kalıntıların bulunduğu bir bölge genelde "sarmal" olarak adlandırılır, gerekli hidrojen bağlarına sahip olmasına bakılmaksızın. Daha az resmî başka tanımlar da öne sürülmüştür, örneğin kavis ve burulma gibi. Hatta bazı yapısal biyologlar atomik çözünürlüklü yeni bir yapıyı çözerken ikincil yapıları "göz kararı" tayin edip bu tayinlerini ilgili PDB dosyasına kaydederler.
Bir nükleik asidin ikincil yapısı bir molekül içindeki baz eşleşme etkileşimlerine değinir. Biyoloji RNA'ların ikincil yapısı genelde sap ve ilmiklere ayrıştırılabilir. Genelde bu elemanlar veya onların bileşimleri ayrıca alt gruplara sınıflandırılabilir, dörtlü dönüş (tetraloop), sahte düğüm (pseudoknot) ve sap-ilmik (stem-loop) gibi. Biyolojik RNA'larda işlevsel öneme sahip olan pek çok ikincil yapı elemanı vardır; bazı örnekler Rho'dan bağımsız sonlandırıcı sap-ilmikler ve tRNA yoncasıdır. RNA moleküllerinin ikincil yapısı deneysel ve berimsel yöntemlerle belirlenir.
Üçüncül yapı
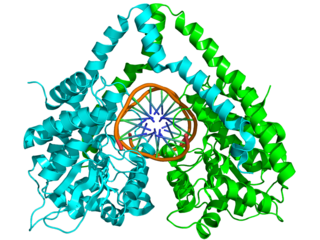
Biyokimya ve moleküler biyolojide, bir protein veya başka bir makromolekülün üçüncül yapısı, onun, atomik koordinatlarla tanımlanmış, üç boyutlu yapısıdır.[6] Protein ve nükleik asitler çeşitli işlevlere sahiptir, moleküler tanımadan katalize kadar uzanan. Bu fonksiyonlar kesin bir üç boyultu yapı gerektirir. Bu yapılar çeşitlilik gösterseler ve karmaşık görünseler de, tekrar tekrar görülen, kolaylıkla tanınabilen üçüncül yapı motiflerinden oluşurlar, bu motifler moleküler yapı blokları olarak görev yaparlar. Üçüncül yapı büyük ölçüde biyomolekülün birincil yapısı tarafından belirlenir. Birincil yapıdan üçüncül yapının öngörülmesi genelde yapı öngörüsü olarak adlandırılır.
Dördüncül yapı
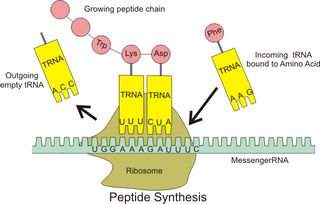
Biyokimyada dördüncül yapı, birden çok katlanmış proteinin çok altbirimli bir kompleks olarak düzenlenmesidir. Nükleik asitlerde bu terim ender kullanılır ama DNA'nın histonlarla etkileşimi veya kromatin içindeki üst seviyeli yapılanmasını,[7] ribozomdaki ayrı RNA birimleri arasındaki etkileşimleri belirtmek için kullanılabilir.[8][9]
Yapı belirlemesi
Yapı yoklaması (İng. structure probing), biyokimyasal yöntemlerle biyomoleküler yapının çözülme sürecidir.[10] Bu analiz, moleküler yapıya kestirmeye yarayabilecek özellikleri tanımlamaya, moleküler yapı ve işlev arasındaki ilişkiyi anlamaya ve başka biyolojik araştırmalara yarayacak yeni küçük moleküllerin geliştirilmesini sağlar.[11] Yapı analizi çeşitli yöntemlerle yapılabilir, bunlar arasında kimyasal yoklamalar, hidroksil radikal yoklaması, nükleotit analog enterferans haritalaması sayılabilir.
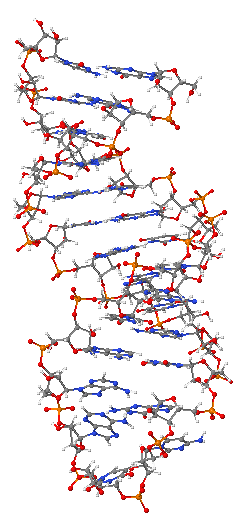
DNA yapısı nükleer manyetik resonans spektroskopisi veya X-ışını kristalografisi ile belirlenebilir. A-DNA'nın X-ışını saçılım örüntüleri hakkındaki ilk rapor, Paterson trasnformuna dayanan bir analize dayalıydı ve yönlendirilmiş DNA lifleri için ancak sınırlı miktarda yapısal bilgi sağlamıştı.[12][13] Daha sonra, 1953'te Wilkins ve arkadaşları alternatif bir analiz yöntemi önerdiler, nemlendirilmiş ve yönlendirilmiş DNA liflerindeki B-DNA yapısının X-ışın kırınım örüntülerinin analizi için Bessel fonksiyonlarını kullandılar.[14] 'B-DNA biçimi' hücre içi şartlarında en sık görülse de,[15] iyi tanımlı bir konformasyon değil, bir DNA konformasyonlar ailesidir.[16] Bunlara karşılık gelen X-ışını kırınım ve saçılım örüntüleri, önemli derecede düzensizlik (>20%) içeren parakristaller için karakteristiktir[17][18] ve dolayıyla standart analizle yapı çözülemez.
Öte yandan, sadece Bessel fonksiyonlarının Fourier transformu ile yapılan standart analiz[19] ve DNA moleküler modeller hâlâ A-DNA ve Z-DNA'nın X-ışını kırınım örüntülerinin analizi için kullanılmaktadır.[20]
Yapı öngörüsü

Biyomolekül yapı öngörüsü bir proteinin üç boyutlu yapısının onun amino asit dizisinden veya bir nükleik asidinkinin onun nükleotit dizisinden öngörülmesidir. Bir diğer deyişle, ikincil ve üçüncül yapıların birincil yapıdan öngörüsüdür. Yapı öngörüsü, biyomoleküler tasarımın tersi olan süreçtir.
Protein yapı öngörüsü biyoenformatik ve teorik kimyanın ulaşmaya çalıştığı en önemli hedeflerden biridir. Protein yapı öngörüsü tıpta (örneğin ilaç tasarımında) ve biyoteknolojide (örneğin yeni enzimlerin tasarımında) çok büyük öneme sahiptir. Güncel yöntemlerin performansı iki yılda düzenlenen CASP yarışmasından değerlendirilir.
RNA yapı öngörü problemi konusu da önemli bir biyoenformatik araştırma konusudur. RNA durumunda nihai yapı molekül içi baz eşleşme etkileşimleri tarafından belirlenir. Bu yüzden farklı nükleotit dizileri olan RNA molekülleri eğer aynı şekilde baz eşleşmesi yapabilirlerse aynı yapıya sahip olabilirler.
Küçük nüklik asit moleküllerinin ikincil yapıları büyük oranda hidrojen bağları ve baz istiflenmesi gibi kuvvetli, lokal etkileşimler tarafından belirlenir. Bu tür etkileşimlerin serbest enerjilerinin toplamı alınarak, genelde en yakın komşu modeli ile, belli bir yapının stabilitesine yaklaşık karşılık gelen bir değer elde edilebilir.[21] En düşük enerji yapısının bulmanın en basit yolu, tüm olası yapıları üretip bunların enerjilerini hesaplamaktır, ama bir diziye ait olası yapıların sayısı, nükleik asidin uzunluğu ile üssel olarak artar.[22] Uzun moleküller için, olasıl ikincil yapıların sayı muazzamdır.[21]
Dizi kovaryasyon (eşdeğişim) yöntemleri, çok sayıda homolog RNA dizilerinden (farklı ama evrimsel olarak ilişkili RNA dizilerinden) oluşan bir veri grubunun varlığına dayanır. Bu yöntemler bireysel nükleotit pozisyonlarının evrim sırasında beraberce değişmeleri irdelenir. Birbirinden uzak iki konumdaki bazların biribiriyle baz eşleşmesi yapacak şekilde beraber değişmesi, bunlar arasındaki baz eşleşmesinin yapısal bakımdan gerekli olduğuna işaret eder. Yalancı düğüm (pseudoknot) öngörüsünün NP-tam olduğu gösterilmiştir.[23]
Tasarım
Biyomoleküler tasarım yapı öngörüsünün tersi sayılabilir. Yapı öngörüsünde, bilinen bir diziden yapı öngörülür, nükleik asit veya protein tasarımında arzu edilen yapıyı meydana getirecek bir dizi elde edilir.
Diğer biyomoleküller
Diğer biyomoleküller de, olisakkaritler ve lipitler gibi, biyolojik öneme sahip, üst düzey yapıya sahip olabilir.
Ayrıca bakınız
- Kodlamayan RNA
- RNA yapı öngörü yazılımları listesi
- Nükleik asit simülasyon yazılımları listesi
Kaynakça
- ^ Samarsky, DA (1998). "The snoRNA box C/D motif directs nucleolar targeting and also couples snoRNA synthesis and localization". EMBO. 17 (13). ss. 3747-3757. doi:10.1093/emboj/17.13.3747. PMC 1170710 $2. PMID 9649444.
- ^ Ganot P, Caizergues-Ferrer M, Kiss T (1997). "The family of box ACA small nucleolar RNAs is defined by an evolutionarily conserved secondary structure and ubiquitous sequence elements essential for RNA accumulation". Genes Dev. 11 (7). ss. 941-56. doi:10.1101/gad.11.7.941. PMID 9106664.
- ^ Shine J, Dalgarno L (1975). "Determinant of cistron specificity in bacterial ribosomes". Nature. 254 (5495). ss. 34-8. doi:10.1038/254034a0. PMID 803646.
- ^ Kozak M (Ekim 1987). "An analysis of 5'-noncoding sequences from 699 vertebrate messenger RNAs". Nucleic Acids Res. 15 (20). ss. 8125-8148. doi:10.1093/nar/15.20.8125. PMC 306349 $2. PMID 3313277. 15 Eylül 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 30 Haziran 2011.
- ^ Bogenhagen DF, Brown DD (1981). "Nucleotide sequences in Xenopus 5S DNA required for transcription termination". Cell. 24 (1). ss. 261-70. doi:10.1016/0092-8674(81)90522-5. PMID 6263489.
- ^ IUPAC, Compendium of Chemical Terminology, 2nd ed. (the "Gold Book") (1997). Online corrected version: (2006–) "tertiary structure 19 Mayıs 2011 tarihinde Wayback Machine sitesinde arşivlendi."
- ^ Sipski, M. Leonide; Wagner, Thomas E. (1977). "Probing DNA quaternary ordering with circular dichroism spectroscopy: Studies of equine sperm chromosomal fibers". Biopolymers. 16 (3). ss. 573-82. doi:10.1002/bip.1977.360160308. PMID 843604.
- ^ Noller, H F (1984). "Structure of Ribosomal RNA". Annual Review of Biochemistry. Cilt 53. ss. 119-62. doi:10.1146/annurev.bi.53.070184.001003. PMID 6206780.
- ^ Nissen, P.; Ippolito, JA; Ban, N; Moore, PB; Steitz, TA (2001). "RNA tertiary interactions in the large ribosomal subunit: The A-minor motif". Proceedings of the National Academy of Sciences. 98 (9). ss. 4899-903. doi:10.1073/pnas.081082398. PMC 33135 $2. PMID 11296253.
- ^ Teunissen AWM (1979). RNA Structure Probing: Biochemical structure analysis of autoimmune-related RNA molecules. ss. 1-27. ISBN 9090132341.
- ^ Pace NR, Thomas BC, Woese CR (1999). Probing RNA Structure, Function, and History by Comparative Analysis. Cold Spring Harbor Laboratory Press. ss. 113-117. ISBN 0879695897.
- ^ Franklin, R.E. and Gosling, R.G. received 6 March 1953. Acta Cryst. (1953). 6, 673: The Structure of Sodium Thymonucleate Fibres I. The Influence of Water Content.; also Acta Cryst. 6, 678: The Structure of Sodium Thymonucleate Fibres II. The Cylindrically Symmetrical Patterson Function.
- ^ Franklin, Rosalind; Gosling, RG (1953). "Molecular Configuration in Sodium Thymonucleate. Franklin R. and Gosling R.G" (PDF). Nature. 171 (4356). ss. 740-741. doi:10.1038/171740a0. PMID 13054694. 3 Ocak 2011 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 30 Haziran 2011.
- ^ Wilkins M.H.F., A.R. Stokes A.R. & Wilson, H.R. (1953). "Molecular Structure of Deoxypentose Nucleic Acids" (PDF). Nature. 171 (4356). ss. 738-740. doi:10.1038/171738a0. PMID 13054693. 13 Mayıs 2011 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 30 Haziran 2011.
- ^ Leslie AG, Arnott S, Chandrasekaran R, Ratliff RL (1980). "Polymorphism of DNA double helices". J. Mol. Biol. 143 (1). ss. 49-72. doi:10.1016/0022-2836(80)90124-2. PMID 7441761.
- ^ Baianu, I.C. (1980). "Structural Order and Partial Disorder in Biological systems". Bull. Math. Biol. 42 (4). ss. 464-468. doi:10.1016/0022-2836(80)90124-2. PMID 7441761.
- ^ Hosemann R., Bagchi R.N., Direct analysis of diffraction by matter, North-Holland Publs., Amsterdam – New York, 1962
- ^ Baianu I.C., X-ray scattering by partially disordered membrane systems, Acta Cryst. A, 34 (1978), 751–753.
- ^ Bessel functions and diffraction by helical structures[] (İngilizce)
- ^ X-Ray Diffraction Patterns of Double-Helical Deoxyribonucleic Acid (DNA) Crystals 24 Temmuz 2009 tarihinde Wayback Machine sitesinde arşivlendi. (İngilizce)
- ^ a b Mathews, D.H. Revolutions in RNA secondary structure prediction. J. Mol. Biol 359, 526-532(2006).
- ^ Zuker, M., Sankoff, D. (1984) RNA secondary structures and their prediction. Bull. Math. Biol. 46,591–621.
- ^ Lyngsø RB, Pedersen CN. (2000). RNA pseudoknot prediction in energy-based models. J Comput Biol 7(3-4): 409-427.