Bakteriyofaj

Bakteriyofaj (Antik Yunanca βακτήριον baktérion bakteri ve φαγεῖν phageín, 'yemek' fiilinden türetme), bakterileri enfekte eden bir virüstür. Terim genelde kısaltılmış hali olan faj olarak kullanılır.
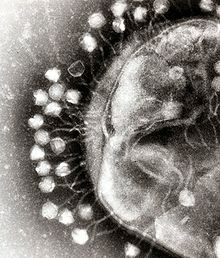
Ökaryotları (hayvan, bitki ve mantarları) enfekte eden virüsler gibi fajlarda da büyük bir yapısal ve işlevsel çeşitlilik vardır. Tipik olarak proteinden oluşan bir kabuk ve içinde yer alan genetik malzemeden oluşurlar. Genetik malzeme DNA veya RNA olabilir, ama genelde 5 - 500 kilo baz çifti uzunluğunda çift sarmallı DNA'dan oluşur. Bakteriyofajlar genelde 20 ila 200 nm arası büyüklükte olurlar.
Fajlar her yerde mevcutturlar ve bakterilerin yaşadığı ortamlarda, örneğin toprakta veya hayvan bağırsaklarında bulunabilirler. Faj ve diğer virüslerin en yoğun doğal kaynaklarından biri deniz suyudur. Deniz yüzeyinde mililitrede 109 etkin faj taneciği (virion) bulunmuştur ve deniz bakterilerinin %70'i fajlar tarafından enfekte olmuş olabilirler.[1]
Tarihçe
1913'te Büyük Britanyalı bakteriyolog Frederick Twort bakterileri enfekte edip öldüren bir etmen keşfetmiş ama konuyu daha fazla takip etmemiştir. Fransız-Kanadalı mikrobiyolog Felix d'Hérelle 3 Eylül 1917'de "dizanteri basilinin düşmanının, görünmez bir mikrobunu" keşfettiğini açıklayıp ona bakteriyofaj adını verdi.
Çoğalması
Bakteriyofajların litik veya lizogenik hayat döngüleri olabilir, bazılarında her ikisi de olur. T4 fajı gibi öldürücü fajlarda görülen litik döngüde virionun çoğalmasının hemen ardından konak hücre parçalanır ve ölür. Hücre ölür ölmez virionların kendilerine yeni bir konak bulmaları gerekir.
Lizogenik döngü, buna tezat olarak, konak hücrenin parçalanmasına neden olmaz. Lizogenik olabilen fajlara ılımlı fajlar (temperate phage) denir. Viral genom konak genoma dahil olur ve oldukça zararsız bir şekilde onunla beraber eşlenir. Konak hücrenin sağlığı yerinde olduğu sürece virüs sessiz bir şekilde varlığını sürdürür, ama konağın şartları bozulursa, örneğin besin kaynaklarının tükenmesi durumunda, endojen fajlar (profaj olarak adlandırılırlar) etkinleşirler. Bir çoğalma süreci başlar, sonucunda konak hücre parçalanır. İlginç bir şekilde lizogenik döngü konak hücrenin çoğalmasına izin verdiği için hücrenin yavrularında da virüs varlığını devam ettirir.
Bazen profajlar inaktif oldukları dönemde bakteri genomuna yeni işlevler kazandırarak konak bakteriye fayda sağlarlar, bu olguya lizogenik dönüşüm (lysogenic conversion) denir. Bunun iyi bilinen bir örneği Vibrio cholerae'nın zararsız bir suşunun bir faj tarafından enfekte edilerek kolera hastalığı etmenine dönüşümüdür.
Bağlanma ve giriş

Konak hücreye girmek için bakteryofajlar bakterinin yüzeyindeki özgül reseptörlere bağlanırlar, bunlar arasında lipopolisakkaritler, teikoik asitler, proteinler sayılabilir. Bu nedenle bir bakteryofaj ancak bağlanabileceği reseptörler taşıyan bakterileri enfekte edebilirler. Faj virionları kendi kendilerine hareket etmedikleri için kendi reseptörleriyle solüsyondayken rassal olarak buluşup bağlanırlar.
Karmaşık bakteryofajlar, örneğin T-çift fajları, genetik malzemelerini hücrenin içine enjekte etmek için şırınga benzeri bir hareket kullanırlar. Uygun reseptörle temas kurduktan sonra kuyruk lifleri taban plakasını hücre yüzeyine yaklaştırırlar. İyice bağlandıktan sonra, kuyruk büzülür, bu da genetik malzemenin dışarı itilmesine neden olur. Bazı fajlar nükleik asiti hücre zarından içeri iter, bazıları hücre yüzeyine birakır. Başka yöntemlerle genetik malzemelerini içeri sokan bakteriyofajlar da vardır.
Protein ve nükleik asit sentezi
Kısa süre, bazen dakikalar içinde, bakteri ribozomları viral mRNA'nın proteine çevirimine (translasyonuna) başlarlar. RNA-fajlarında RNA-replikaz bu sürecin başlarında sentezlenir. Erken sentezlenen proteinler ve virionla gelen bazı proteinler bakterinin RNA polimerazını modifiye edip onun viral mRNA'yı tercihen çevirmesine neden olabilirler. Konağın kendi protein ve nükleik asit sentezi de bozularak viral ürünlerin sentezine yönlendirilir. Bu ürünler ya hücreyi parçalamaya yarayacaklar, ya yeni virionların oluşmasına yardımcı olacaklar veya yeni virionları oluşturacaklardır.
Virion oluşumu
T4 fajları durumunda yeni fajların inşası özel yardımcı molekülleri gerektiren karmaşık bir süreçtir. Önce taban plakası oluşur, kuyruk onun üzerinde büyür. Kafa kapsidi, ayrı olarak oluşup kendiliğinden kuyruk ile birleşir. Henüz bilinmeyen bir şekilde DNA kafanın içine sıkı bir şekilde yerini alır. Bütün süreç yaklaşık 15 dakika alır.
Virionların salınımı
Fajlar ya hücre parçalanması (lizis) veya salgılanma yoluyla salınırlar. T4 fajları durumunda, hücre içine girmelerinden 20 dakikadan biraz sonra hücre parçalanması yoluyla sayıları 300'ü bulabilen faj salınır. Bunun gerçekleşmesi, hücre duvarındaki peptidoglikanı parçalayan endolizin adlı enzim sayesinde olur. Bazı virüsler ise parazite dönüşüp konak hücrenin sürekli olarak yeni virüs tanecikleri salgılamasına neden olabilirler. Yeni virionlar hücre zarından tomurcuklanarak koparlar, beraberlerinde hücre zarının bir kısmını da götüren bu fajlar örtülü virüs olarak ortama salınırlar. Salınan virionların her biri yeni bir bakteriyi enfekte edebilir.
Faj terapisi
Keşiflerinin ardından fajlar anti-bakteriyel etmen olarak denenmişlerdir. Ancak antibiyotikler keşfedilince bunların fajlardan daha kullanışlı oldukları görülmüştür ve Batı'da faj tedavisi üzerine yapılan araştırmalar bırakılmıştır. Buna karşın Sovyetler Birliği'nde 1940'lardan beri antibiyotiklere alternatif olarak kullanımı devam etmiştir.
Bakteri suşlarında doğal seleksiyon yoluyla antibiyotik direncinin oluşması bazı tıbbi araştırmacıları faj tedavisini antibiyotik tedavisine bir alternatif olarak tekrar değerlendirmeye sevketmiştir. Antibiyotiklerden farklı olarak fajlar, milyonlarca yıldır süregeldiği gibi, bakterilerle beraber evrimleştikleri için, sürekli bir direncin oluşma olasılığı yok sayılabilir. Ayrıca, etkili bir faj, özgül bakterisini tamamen bitene kadar enfekte etmeye devam edecektir.
Belli bir faj genelde ancak belli bir bakteri tipini enfekte edebildiği için, ki bu birkaç bakteri türü olabileceği gibi bir türün sadece bazı alt türleri de olabilir, bakteri tipinin doğru tanımlandığından emin olmak gerekebilir, bu da 24 saat sürebilir. Faj terapisinin bir diğer avantajı başka bakterilere zarar gelmeyeceğinden dar spektrumlu antibiyotik terapisine benzemesidir. Ancak, sıkça olduğu gibi, birden fazla bakterinin beraberce neden oldukları enfeksiyonlarda bu bir dezavantaj oluşturabilir. Bakteriyofajların bir diğer sorunu vücudun bağışıklık sisteminin saldırısına uğramalarıdır.
Fajlar enfeksiyonla doğrudan temas durumunda etki gösterirler, onun için açık bir yaraya uygulanmaları en iyi sonuç doğurur. Sistemik enfeksiyonlarda bu pratik olarak mümkün değildir. Sovyetler birliğinde diğer tedavilerin çalışmadığı durumlarda gözlenen başarılı sonuçlara rağmen çoğu araştırmacı faj terapisinin tıbbi bir geçerliliğe ulaşacağına şüphe ile bakmaktadır. Faj tedavisinin etkinliğini belirlemek için büyük ölçekli klink testler yapılmamıştır ama antibiyotik dirençli bakteri türlerinin çoğalmasından dolayı bu konuda araştırmalar sürmektedir.
Ağustos 2006'da ABD Gıda ve İlaç İdaresi (Food and Drug Administration) bazı etlerde Listeria monocytogenes bakterisinin öldürülmesi için bakteriyofaj kullanımını onaylamıştır.[2]
Faj terapisinin avantaj ve dezavantajları
Fajların antibiyotiklere göre bazı avantaj ve dezavantajları vardır. (Tablo 1) Bunlar içinde en önemlileri; fajın uygulanma biçiminden bağımsız olarak nerede ihtiyaç varsa oraya göç etmesi ve orada çoğalması, antibiyotik direncinden bağımsız olarak aktivite göstermesi, biyofilm üzerine etkin olması, etki spektrumunun dar olması ve faj terapisi öncesi etken/faj ilişkisinin tanımlanmış olması olarak sıralanabilir.[3]
Faj terapisinin güvenilirliği: Günümüze kadar yapılan çalışmalarda faj terapisinin, minör etkileri dışında, hiçbir yan etkisi bildirilmemiştir.[3]
| Bakteriyofajlar | Antibiyotikler | Yorum | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Etki spektrumu oldukça dar ve oldukça spesifiktir. | Etki spektrumu geniştir. | Faj tedavisi öncesi bakterinin tanımlanması ve faj duyarlılığının tespit edilmesi gerekliliği sebebiyle, yüksek spesifikte fajın dezavantajları arasında sayılabilir. Antibiyotiklerde etki spektrumu geniş olabilir ancak mikrobiyal etkiyi büyük ölçüde etkiler. | |||||||||||||||||||||||||||||||||
| Enfeksiyonun olduğu yerde replike olur ve nerede ihtiyaç varsa orada çoğalır. | Vücutta metabolize edilir ve eliminasyona uğrar. | Fajlar geometrik şekilde çoğaldıklarından enfeksiyonun olduğu yere az sayıda enjekte edilmesi yeterli olacaktır. Antibiyotiklerin enfeksiyon bölgesinde her zaman konsantre olmaması sebebiyle yüksek doz ve belirli aralıklarla uygulanması gerekir. | |||||||||||||||||||||||||||||||||
| Hızlı bakterisit etki gösterir. | Bakterisit / Bakteriyostatik etki gösterebilir. | Hızlı bakterisit etki sebebiyle fajlar akut vakalarda hızlı etki gösterebilirler. Bakteriyostatik antibiyotik etkileri bakterilerde direnç gelişimine neden olabilir. | |||||||||||||||||||||||||||||||||
| Şu ana kadar gözlemlenmiş bir yan etkisi bulunmamaktadır. | Birçok yan etkisi vardır. | Faj tarafından parçalanan bakterilerden salınan endotoksinlere bağlı olarak çok az yan etki bildirilmiştir. | |||||||||||||||||||||||||||||||||
| Faja dirençli bakteriler aynı görevdeki fajlara karşı duyarlı kalabilir. | Antibiyotik direnci sadece hedeflenen bakteriyle sınırlı kalmaz. | "Doğada bir bakteri varsa, onu enfekte eden faj da mutlaka vardır." varsayımı dolayısıyla, bir faja karşı dirençli olan bakteriler için yeni fajlar kolaylıkla izole edilebilinir. Antibiyotik direnci özellikle hareketli genetik araçlarla aktarılabilinir. | |||||||||||||||||||||||||||||||||
| Faj seçimi hızlı bir faaliyet olup günler, haftalar içinde yapılabilir. Oldukça düşük maliyetlidir. | Yeni antibiyotik geliştirilmesi oldukça uzun zaman ve maliyet ister. | Doğada istenen konağa karşı kullanılacak fajın izolasyonu oldukça kolay ve düşük maliyetlidir. Fakat doğru fajın seçilip terapide kullanılacak hale getirilebilinmesi için uzmana ihtiyaç vardır. Bakterinin kültürü ne kadar zorsa faj izolasyonu da o derecede zordur. | |||||||||||||||||||||||||||||||||
| İnsan vücudundaki faj aktivitesi azaltıcı etkenlere maruz kalabilir. | Aktivite azaltıcı etkenlerden korunmak için standardizasyonları oldukça iyi yapılmıştır. | Antibiyotiklerin kullanım kolaylığının yanında fajların bu özelliği dezavantaj olarak kalmaktadır. Ancak ileri teknolojilerle faj preparasyonlarının kapsül haline getirilmesi ve azaltıcı etkenlerden korunması etkili yöntemler arasında gösterilmektedir. | |||||||||||||||||||||||||||||||||
Faj direnci: Bakteriyofaj ve konağı olan bakterilerin (Devam edecek.)
Model bakteriyofajlar
Aşağıda ayrıntılı olarak üzerinde çalışılmış olan bakteriyofajların bir listesi bulunmaktadır:
- λ faj
- T4 fajı
- T7 fajı
- R17 fajı
- M13 fajı
- MS2 fajı
- P1 fajı
- P2 fajı
- N4 fajı
- Φ6 fajı
- Ф29 fajı
Kaynakça
- Häusler, T. (2006) "Viruses vs. Superbugs", Macmillan(İngilizce)
- Evergreen State College material on bacteriophages7 Aralık 2006 tarihinde Wayback Machine sitesinde arşivlendi.(İngilizce)
- Phage.org general information on bacteriophages(İngilizce)
- Pittsburgh Bacteriophage Institute (İngilizce)
- ^ Prescott, L. (1993). Microbiology, Wm. C. Brown Publishers, ISBN 0-697-01372-3
- ^ "FDA Approval of Listeria-specific Bacteriophage Preparation on Ready-to-Eat (RTE) Meat and Poultry Products" [1] 14 Mayıs 2007 tarihinde Wayback Machine sitesinde arşivlendi. ABD Gıda ve İlaç idaresi (FDA) duyurusu (İngilizce)
- ^ a b "Arşivlenmiş kopya" (PDF). 1 Kasım 2021 tarihinde kaynağından arşivlendi (PDF). Erişim tarihi: 1 Kasım 2021.
Dış bağlantılar
| Bileşenler |
|  |
|---|---|---|
| Viral hayat döngüsü |
| |
| Genetik |
| |
| Konak |
| |
| Etkileri |
| |
| Belirtiler ve Testler | ||
| Diğerleri |
| |










