İlaç metabolizması
İlaç metabolizması, ilaçların canlı organizmalar tarafından, genellikle özel enzimatik sistemler aracılığıyla metabolik olarak parçalanmasıdır. Daha genel olarak, ksenobiyotik metabolizması (Yunanca xenos "yabancı" ve biotic "canlılarla ilgili"), herhangi bir ilaç veya zehir gibi bir organizmanın normal biyokimyasına yabancı bileşikler olan ksenobiyotiklerin kimyasal yapısını değiştiren metabolik yollar kümesidir. Bu yollar, tüm büyük organizma gruplarında bulunan bir biyotransformasyon şeklidir ve antik kökenli olduğu düşünülmektedir. Bu reaksiyonlar genellikle zehirli bileşiklerin detoksifikasyonunu sağlar (bazı durumlarda ksenobiyotik metabolizmasındaki ara ürünler toksik etkilere neden olabilir). İlaç metabolizmasının incelenmesine farmakokinetik denir.
Farmasötik ilaçların metabolizması, farmakoloji ve tıbbın önemli bir yönüdür. Örneğin, metabolizma hızı bir ilacın farmakolojik etkisinin süresini ve yoğunluğunu belirler. İlaç metabolizması ayrıca bulaşıcı hastalıklarda ve kanser kemoterapisinde çoklu ilaç direncini etkiler ve bazı ilaçların ksenobiyotik metabolizmasında yer alan enzimlerin substratları veya inhibitörleri olarak etkileri, tehlikeli ilaç etkileşimlerinin yaygın bir nedenidir. Bu yollar çevre biliminde de önemlidir; mikroorganizmaların ksenobiyotik metabolizması, bir kirleticinin biyoremediasyon sırasında parçalanıp parçalanmayacağını veya çevrede kalıcı olup olmayacağını belirler. Ksenobiyotik metabolizma enzimleri, özellikle de glutatyon S-transferazlar, pestisitlere ve herbisitlere karşı direnç üretebildikleri için tarımda da önemlidir.
İlaç metabolizması üç faza ayrılır. Faz I'de sitokrom P450 oksidazlar gibi enzimler ksenobiyotiklere reaktif veya polar gruplar ekler. Bu modifiye bileşikler daha sonra faz II reaksiyonlarında polar bileşiklere konjuge edilir. Bu reaksiyonlar glutatyon S-transferazlar gibi transferaz enzimleri tarafından katalize edilir. Son olarak, faz III'te, konjuge ksenobiyotikler efluks taşıyıcıları tarafından tanınmadan ve hücrelerden dışarı pompalanmadan önce daha fazla işlenebilir. İlaç metabolizması genellikle lipofilik bileşikleri daha kolay atılabilen hidrofilik ürünlere dönüştürür.[]
Geçirgenlik bariyerleri ve detoksifikasyon
Bir organizmanın maruz kaldığı kesin bileşikler büyük ölçüde öngörülemez ve zaman içinde büyük farklılıklar gösterebilir; bunlar ksenobiyotik toksik stresin başlıca özellikleridir.[1] Ksenobiyotik detoksifikasyon sistemlerinin karşılaştığı en büyük zorluk, normal metabolizmada yer alan karmaşık kimyasal karışımından neredeyse sınırsız sayıda ksenobiyotik bileşiği çıkarabilmeleri gerektiğidir. Bu sorunu ele almak için geliştirilen çözüm, fiziksel engeller ve düşük özgüllüklü enzimatik sistemlerin zarif bir kombinasyonudur.
Tüm organizmalar iç ortamlarına erişimi kontrol etmek için hücre zarlarını hidrofobik geçirgenlik bariyerleri olarak kullanır. Polar bileşikler bu hücre zarları boyunca yayılamaz ve yararlı moleküllerin alımına, hücre dışı karışımdan substratları özel olarak seçen taşıma proteinleri aracılık eder. Bu seçici alım, çoğu hidrofilik molekülün herhangi bir spesifik taşıyıcı tarafından tanınmadığı için hücrelere giremeyeceği anlamına gelir.[2] Buna karşılık, hidrofobik bileşiklerin bu bariyerler boyunca difüzyonu kontrol edilemez ve bu nedenle organizmalar membran bariyerlerini kullanarak lipitte çözünen ksenobiyotikleri dışlayamaz.
Bununla birlikte, bir geçirgenlik bariyerinin varlığı, organizmaların membran geçirgen ksenobiyotiklerde ortak olan hidrofobiklikten yararlanan detoksifikasyon sistemleri geliştirebildikleri anlamına gelir. Dolayısıyla bu sistemler, polar olmayan hemen her bileşiği metabolize edecek kadar geniş substrat spesifikliklerine sahip olarak spesifiklik sorununu çözmektedir.[1] Faydalı metabolitler polar olduklarından ve genel olarak bir veya daha fazla yüklü grup içerdiklerinden hariç tutulurlar.
Normal metabolizmanın reaktif yan ürünlerinin detoksifikasyonu yukarıda özetlenen sistemler tarafından gerçekleştirilemez, çünkü bu türler normal hücresel bileşenlerden türetilir ve genellikle polar özelliklerini paylaşırlar. Bununla birlikte, bu bileşikler sayıca az olduğundan, spesifik enzimler bunları tanıyabilir ve uzaklaştırabilir. Bu spesifik detoksifikasyon sistemlerine örnek olarak reaktif aldehit metilglioksal'ı ortadan kaldıran glioksalaz sistemi[3] ve reaktif oksijen türlerini ortadan kaldıran çeşitli antioksidan sistemler verilebilir.[4]
Detoksifikasyon aşamaları
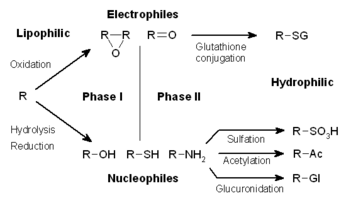
Ksenobiyotiklerin metabolizması genellikle üç aşamaya ayrılır: modifikasyon, konjugasyon ve atılım. Bu reaksiyonlar, ksenobiyotikleri detoksifiye etmek ve hücrelerden uzaklaştırmak için birlikte hareket eder.
Faz I - modifikasyon
Faz I'de, çeşitli enzimler substratlarına reaktif ve polar gruplar eklemek için hareket eder. En yaygın modifikasyonlardan biri sitokrom P450'ye bağlı karışık fonksiyonlu oksidaz sistemi tarafından katalize edilen hidroksilasyondur. Bu enzim kompleksleri, aktive olmamış hidrokarbonlara bir oksijen atomu eklemek için hareket eder, bu da hidroksil gruplarının eklenmesine veya substratların N-, O- ve S-dealkilasyonuna neden olabilir.[5] P450 oksidazların reaksiyon mekanizması, aşağıdaki şemaya göre sitokroma bağlı oksijenin indirgenmesi ve oldukça reaktif bir oksiferril türünün üretilmesi yoluyla ilerler:[6]
- O2 + NADPH + H+ + RH → NADP+ + H2O + ROH
Faz I reaksiyonları (sentetik olmayan reaksiyonlar olarak da adlandırılır) oksidasyon, redüksiyon, hidroliz, siklizasyon, desiklizasyon ve oksijen eklenmesi veya hidrojen çıkarılması yoluyla meydana gelebilir ve genellikle karaciğerde karışık fonksiyonlu oksidazlar tarafından gerçekleştirilir. Bu oksidatif reaksiyonlar tipik olarak bir sitokrom P450 monooksijenaz (genellikle CYP olarak kısaltılır), NADPH ve oksijen içerir. Metabolizmaları için bu yöntemi kullanan farmasötik ilaç sınıfları arasında fenotiyazinler, parasetamol ve steroidler bulunur. Faz I reaksiyonlarının metabolitleri yeterince polar ise, bu noktada kolayca atılabilirler. Bununla birlikte, birçok faz I ürünü hızlı bir şekilde elimine edilmez ve endojen bir substratın yeni dahil edilen fonksiyonel grupla birleşerek oldukça polar bir konjugat oluşturduğu müteakip bir reaksiyona girer.
Yaygın bir Faz I oksidasyonu, bir C-H bağının bir C-OH'ye dönüştürülmesini içerir. Bu reaksiyon bazen farmakolojik olarak inaktif bir bileşiği (bir ön ilaç) farmakolojik olarak aktif bir bileşiğe dönüştürür. Aynı şekilde, Faz I toksik olmayan bir molekülü zehirli bir moleküle dönüştürebilir (toksikasyon). Midedeki basit hidroliz normalde zararsız bir reaksiyondur, ancak istisnalar da vardır. Örneğin, faz I metabolizması asetonitrili HOCH2CN'ye dönüştürür, bu da hızla formaldehit ve hidrojen siyanüre ayrışır.[7]
İlaç adaylarının Faz I metabolizması, enzim olmayan katalizörler kullanılarak laboratuvarda simüle edilebilir.[8] Bu biyomimetik reaksiyon örneği, genellikle Faz I metabolitlerini içeren ürünler verme eğilimindedir. Örnek olarak, farmasötik trimebutinin ana metaboliti olan desmetiltrimebutin (nor-trimebutin), piyasada bulunan ilacın in vitro oksidasyonu ile verimli bir şekilde üretilebilir. Bir N-metil grubunun hidroksilasyonu bir formaldehit molekülünün dışarı atılmasına yol açarken, O-metil gruplarının oksidasyonu daha az ölçüde gerçekleşir.
Oksidasyon
- Sitokrom P450 monooksijenaz sistemi
- Flavin içeren monooksijenaz sistemi
- Alkol dehidrojenaz ve aldehit dehidrojenaz
- Monoamin oksidaz
- Peroksidazlar tarafından ko-oksidasyon
Redüksiyon
- NADPH-sitokrom P450 redüktaz
Sitokrom P450 redüktaz - NADPH:ferrihemoprotein oksidoredüktaz, NADPH:hemoprotein oksidoredüktaz, NADPH:P450 oksidoredüktaz, P450 redüktaz, POR, CPR, CYPOR olarak da bilinir - FAD ve FMN içeren bir enzim olan NADPH:sitokrom P450 redüktazdan ökaryotik hücrenin mikrozomundaki sitokrom P450'ye elektron transferi için gerekli olan membrana bağlı bir enzimdir POR/P450 sistemindeki elektron akışının genel şeması şöyledir: NADPH → FAD → FMN → P450 → O2
- İndirgenmiş (redükte/demirli) sitokrom P450
İndirgeme (redüksiyon) reaksiyonları sırasında bir kimyasal, bir serbest radikal elektron kazandığı ve hemen ardından bunu oksijene (süperoksit anyonu oluşturmak üzere) kaybettiği boş döngüye girebilir.
Hidroliz
- Esterazlar ve amidazlar
- Epoksit hidrolaz
Faz II - konjugasyon
Sonraki faz II reaksiyonlarında, bu aktive edilmiş ksenobiyotik metabolitler glutatyon (GSH), sülfat, glisin veya glukuronik asit gibi yüklü türlerle konjuge edilir. İlaçlar üzerinde konjugasyon reaksiyonlarının gerçekleştiği bölgeler arasında karboksi (-COOH), hidroksi (-OH), amino (NH2) ve tiyol (-SH) grupları bulunmaktadır.
Konjugasyon reaksiyonlarının ürünleri, genellikle aktif metabolitler üreten Faz I reaksiyonlarının aksine, artan moleküler ağırlığa sahiptir ve substratlarından daha az aktif olma eğilimindedir. Büyük anyonik grupların (GSH gibi) eklenmesi reaktif elektrofilleri detoksifiye eder ve membranlar boyunca difüze olamayan daha polar metabolitler üretir ve bu nedenle aktif olarak taşınabilir.
Bu reaksiyonlar, kombinasyon halinde nükleofilik veya elektrofilik gruplar içeren neredeyse tüm hidrofobik bileşikleri metabolize edebilen geniş spesifiklikli transferazların büyük bir grubu tarafından katalize edilir.[1] Bu grubun en önemli sınıflarından biri glutatyon S-transferazlardır (GST'ler).
| Mekanizma | İlgili enzim | Eşfaktör | Konum | Kaynaklar |
|---|---|---|---|---|
| metilasyon | metiltransferaz | S-adenosil-L-metiyonin | karaciğer, böbrek, akciğer, MSS | [9] |
| sülfasyon | sülfotransferaz | 3'-Fosfoadenosin-5'-fosfosülfat | karaciğer, böbrek, bağırsak | [9] |
| asetilasyon |
| asetil koenzim A | karaciğer, akciğer, dalak, mide mukozası, alyuvarlar, lenfositler | [9] |
| glukuronidasyon | UDP-glukuronoziltransferazlar | UDP-glukuronik asit | karaciğer, böbrek, bağırsak, akciğer, deri, prostat, beyin | [9] |
| glutatyon konjugasyonu | glutatyon S-transferaz | glutatyon | karaciğer, böbrek | [9] |
| glisin konjugasyonu | İki aşamalı süreç:
| glisin | karaciğer, böbrek | [10] |
Faz III - daha fazla modifikasyon ve atılım
Faz II reaksiyonlarından sonra, ksenobiyotik konjugatlar daha fazla metabolize edilebilir. Yaygın bir örnek, glutatyon konjugatlarının asetilsistein (merkapturik asit) konjugatlarına işlenmesidir.[11] Burada glutatyon molekülündeki γ-glutamat ve glisin kalıntıları gama-glutamil transpeptidaz ve dipeptidazlar tarafından uzaklaştırılır. Son adımda, konjugattaki sistein kalıntısı asetillenir.
Konjugatlar ve metabolitleri metabolizmalarının III. fazında hücrelerden atılabilir, anyonik gruplar çoklu ilaç direnci protein (MRP) ailesinin çeşitli membran taşıyıcıları için afinite etiketleri görevi görür.[12] Bu proteinler ATP-bağlayıcı kaset taşıyıcıları ailesinin üyeleridir ve çok çeşitli hidrofobik anyonların ATP'ye bağlı taşınmasını katalize edebilir[13] ve böylece faz II ürünlerini daha fazla metabolize edilebilecekleri veya atılabilecekleri hücre dışı ortama uzaklaştırmak için hareket edebilirler.[14]
Endojen toksinler
Peroksitler ve reaktif aldehitler gibi endojen reaktif metabolitlerin detoksifikasyonu genellikle yukarıda açıklanan sistemle sağlanamaz. Bunun nedeni, bu türlerin normal hücresel bileşenlerden türetilmesi ve genellikle polar özelliklerini paylaşmasıdır. Ancak, bu bileşikler sayıca az olduğundan, enzimatik sistemlerin bunları tanımak ve uzaklaştırmak için spesifik moleküler tanıma kullanması mümkündür. Bu moleküllerin faydalı metabolitlere olan benzerliği, her bir endojen toksin grubunun metabolizması için genellikle farklı detoksifikasyon enzimlerinin gerekli olduğu anlamına gelmektedir. Bu spesifik detoksifikasyon sistemlerine örnek olarak reaktif aldehit metilglioksalın atılmasını sağlayan glioksalaz sistemi ve reaktif oksijen türlerini ortadan kaldıran çeşitli antioksidan sistemler verilebilir.
Bölgeler
Her biyolojik doku ilaçları metabolize etme yeteneğine sahip olsa da - niceliksel olarak - karaciğer hücresinin düz endoplazmik retikulumu ilaç metabolizmasının ana organıdır. Karaciğerin ilaç metabolizmasına katkısından sorumlu faktörler arasında büyük bir organ olması, bağırsakta emilen kimyasallar tarafından perfüze edilen ilk organ olması ve diğer organlara göre çoğu ilaç metabolize edici enzim sisteminin çok yüksek konsantrasyonlarda bulunması yer alır. Bir ilaç portal ven yoluyla hepatik dolaşıma girdiği GI kanalına alınırsa, iyi metabolize olur ve ilk geçiş etkisi gösterdiği söylenir.
İlaç metabolizmasının diğer bölgeleri gastrointestinal sistem, akciğerler, böbrekler ve derinin epitel hücrelerini içerir. Bu bölgeler genellikle lokalize toksisite reaksiyonlarından sorumludur.
İlaç metabolizmasını etkileyen faktörler
Çoğu lipofilik ilacın farmakolojik etki süresi ve yoğunluğu, inaktif ürünlere metabolize edilme hızına göre belirlenir. Sitokrom P450 monooksijenaz sistemi bu konudaki en önemli yoldur. Genel olarak, farmakolojik olarak aktif bir metabolitin metabolizma hızını (örn. enzim indüksiyonu) artıran herhangi bir şey, ilacın etki süresini ve yoğunluğunu azaltacaktır. Bunun tersi de doğrudur (örn. enzim inhibisyonu). Bununla birlikte, bir enzimin bir ön ilacın bir ilaca metabolize edilmesinden sorumlu olduğu durumlarda, enzim indüksiyonu bu dönüşümü hızlandırabilir ve ilaç seviyelerini artırarak potansiyel olarak toksisiteye neden olabilir.
Çeşitli fizyolojik ve patolojik faktörler de ilaç metabolizmasını etkileyebilir. İlaç metabolizmasını etkileyebilecek fizyolojik faktörler arasında yaş, bireysel farklılıklar (örn. farmakogenetik), enterohepatik dolaşım, beslenme, bağırsak florası veya cinsiyet farklılıkları yer almaktadır.
Genel olarak, ilaçlar fetal, yenidoğan ve yaşlı insan ve hayvanlarda yetişkinlere göre daha yavaş metabolize olur.
Genetik varyasyon (polimorfizm) ilaçların etkisindeki değişkenliğin bir kısmını açıklamaktadır. N-asetiltransferazlar (Faz II reaksiyonlarında yer alır) ile bireysel varyasyon, yavaş asetilasyon yapan (yavaş asetilatörler) ve hızlı asetilasyon yapan bir grup insan yaratır ve Kanada nüfusunda kabaca 50:50 oranında bölünür. Yavaş asetilleyiciler doza bağlı toksisiteye daha yatkın olduklarından, bu varyasyonun dramatik sonuçları olabilir.
Sitokrom P450 monooksijenaz sistemi enzimleri de bireyler arasında farklılık gösterebilir ve etnik kökenlerine bağlı olarak insanların %1-30'unda eksiklikler görülür.
İlacın dozu, sıklığı, uygulama yolu, doku dağılımı ve proteine bağlanması metabolizmasını etkiler.
Karaciğer, böbrek veya kalp hastalıkları gibi patolojik faktörler de ilaç metabolizmasını etkileyebilir.
In silico modelleme ve simülasyon yöntemleri, insan deneklerde klinik çalışmalar yapılmadan önce sanal hasta popülasyonlarında ilaç metabolizmasının tahmin edilmesini sağlar.[15] Bu, advers reaksiyon açısından en fazla risk altında olan bireyleri belirlemek için kullanılabilir.
Tarihçe
İnsanların aldıkları maddeleri nasıl dönüştürdüklerine dair çalışmalar on dokuzuncu yüzyılın ortalarında, kimyagerlerin benzaldehit gibi organik kimyasalların insan vücudunda oksitlenebileceğini ve amino asitlere konjuge edilebileceğini keşfetmeleriyle başlamıştır.[16] On dokuzuncu yüzyılın geri kalanında metilasyon, asetilasyon ve sülfonasyon gibi diğer bazı temel detoksifikasyon reaksiyonları keşfedilmiştir.
Yirminci yüzyılın başlarında, çalışmalar bu metabolitlerin üretiminden sorumlu olan enzimlerin ve yolakların araştırılmasına yönelmiştir. Bu alan, Richard Williams'ın 1947 yılında "Detoksifikasyon Mekanizmaları" kitabını yayınlamasıyla ayrı bir çalışma alanı olarak tanımlanmıştır.[17] Bu modern biyokimyasal araştırma, 1961 yılında glutatyon S-transferazların tanımlanmasıyla sonuçlanmış,[18] bunu 1962 yılında sitokrom P450'lerin keşfi[19] ve 1963 yılında ksenobiyotik metabolizmasındaki merkezi rollerinin fark edilmesi izlemiştir.[20][21]
Ayrıca bakınız
Kaynakça
- ^ a b c Jakoby WB, Ziegler DM (December 1990). "The enzymes of detoxication". J. Biol. Chem. 265 (34). ss. 20715-8. doi:10.1016/S0021-9258(17)45272-0. PMID 2249981. 21 Haziran 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Aralık 2012.
- ^ Mizuno N, Niwa T, Yotsumoto Y, Sugiyama Y (September 2003). "Impact of drug transporter studies on drug discovery and development". Pharmacol. Rev. 55 (3). ss. 425-61. doi:10.1124/pr.55.3.1. PMID 12869659.
- ^ Thornalley PJ (July 1990). "The glyoxalase system: new developments towards functional characterization of a metabolic pathway fundamental to biological life". Biochem. J. 269 (1). ss. 1-11. doi:10.1042/bj2690001. PMC 1131522 $2. PMID 2198020.
- ^ Sies H (March 1997). "Oxidative stress: oxidants and antioxidants". Exp. Physiol. 82 (2). ss. 291-5. doi:10.1113/expphysiol.1997.sp004024. PMID 9129943.
- ^ Guengerich FP (June 2001). "Common and uncommon cytochrome P450 reactions related to metabolism and chemical toxicity". Chem. Res. Toxicol. 14 (6). ss. 611-50. doi:10.1021/tx0002583. PMID 11409933.
- ^ Schlichting I, Berendzen J, Chu K, Stock AM, Maves SA, Benson DE, Sweet RM, Ringe D, Petsko GA, Sligar SG (March 2000). "The catalytic pathway of cytochrome p450cam at atomic resolution". Science. 287 (5458). ss. 1615-22. Bibcode:2000Sci...287.1615S. doi:10.1126/science.287.5458.1615. PMID 10698731.
- ^ "Acetonitrile (EHC 154, 1993)". www.inchem.org. 22 Mayıs 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 3 Mayıs 2017.
- ^ Akagah B, Lormier AT, Fournet A, Figadère B (December 2008). "Oxidation of antiparasitic 2-substituted quinolines using metalloporphyrin catalysts: scale-up of a biomimetic reaction for metabolite production of drug candidates". Org. Biomol. Chem. 6 (24). ss. 4494-7. doi:10.1039/b815963g. PMID 19039354.
- ^ a b c d e Liston HL, Markowitz JS, DeVane CL (October 2001). "Drug glucuronidation in clinical psychopharmacology". J Clin Psychopharmacol. 21 (5). ss. 500-15. doi:10.1097/00004714-200110000-00008. PMID 11593076.
- ^ Badenhorst CP, van der Sluis R, Erasmus E, van Dijk AA (September 2013). "Glycine conjugation: importance in metabolism, the role of glycine N-acyltransferase, and factors that influence interindividual variation". Expert Opinion on Drug Metabolism & Toxicology. 9 (9). ss. 1139-1153. doi:10.1517/17425255.2013.796929. PMID 23650932.
Glycine conjugation of mitochondrial acyl-CoAs, catalyzed by glycine N-acyltransferase (GLYAT, E.C. 2.3.1.13), is an important metabolic pathway responsible for maintaining adequate levels of free coenzyme A (CoASH). However, because of the small number of pharmaceutical drugs that are conjugated to glycine, the pathway has not yet been characterized in detail. Here, we review the causes and possible consequences of interindividual variation in the glycine conjugation pathway. ...
Figure 1. Glycine conjugation of benzoic acid. The glycine conjugation pathway consists of two steps. First benzoate is ligated to CoASH to form the high-energy benzoyl-CoA thioester. This reaction is catalyzed by the HXM-A and HXM-B medium-chain acid:CoA ligases and requires energy in the form of ATP. ... The benzoyl-CoA is then conjugated to glycine by GLYAT to form hippuric acid, releasing CoASH. In addition to the factors listed in the boxes, the levels of ATP, CoASH, and glycine may influence the overall rate of the glycine conjugation pathway. - ^ Boyland E, Chasseaud LF (1969). "The role of glutathione and glutathione S-transferases in mercapturic acid biosynthesis". Adv. Enzymol. Relat. Areas Mol. Biol. Advances in Enzymology – and Related Areas of Molecular Biology. Cilt 32. ss. 173-219. doi:10.1002/9780470122778.ch5. ISBN 9780470122778. PMID 4892500.
- ^ Homolya L, Váradi A, Sarkadi B (2003). "Multidrug resistance-associated proteins: Export pumps for conjugates with glutathione, glucuronate or sulfate". BioFactors. 17 (1–4). ss. 103-14. doi:10.1002/biof.5520170111. PMID 12897433.
- ^ König J, Nies AT, Cui Y, Leier I, Keppler D (December 1999). "Conjugate export pumps of the multidrug resistance protein (MRP) family: localization, substrate specificity, and MRP2-mediated drug resistance". Biochim. Biophys. Acta. 1461 (2). ss. 377-94. doi:10.1016/S0005-2736(99)00169-8. PMID 10581368.
- ^ Commandeur JN, Stijntjes GJ, Vermeulen NP (June 1995). "Enzymes and transport systems involved in the formation and disposition of glutathione S-conjugates. Role in bioactivation and detoxication mechanisms of xenobiotics". Pharmacol. Rev. 47 (2). ss. 271-330. PMID 7568330.
- ^ Rostami-Hodjegan A, Tucker GT (February 2007). "Simulation and prediction of in vivo drug metabolism in human populations from in vitro data". Nat Rev Drug Discov. 6 (2). ss. 140-8. doi:10.1038/nrd2173. PMID 17268485.
- ^ Murphy PJ (June 2001). "Xenobiotic metabolism: a look from the past to the future". Drug Metab. Dispos. 29 (6). ss. 779-80. PMID 11353742. 21 Haziran 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Aralık 2012.
- ^ Neuberger A, Smith RL (1983). "Richard Tecwyn Williams: the man, his work, his impact". Drug Metab. Rev. 14 (3). ss. 559-607. doi:10.3109/03602538308991399. PMID 6347595.
- ^ Booth J, Boyland E, Sims P (June 1961). "An enzyme from rat liver catalysing conjugations with glutathione". Biochem. J. 79 (3). ss. 516-24. doi:10.1042/bj0790516. PMC 1205680 $2. PMID 16748905.
- ^ Omura T, Sato R (April 1962). "A new cytochrome in liver microsomes". J. Biol. Chem. 237 (4). ss. 1375-6. doi:10.1016/S0021-9258(18)60338-2. PMID 14482007. 21 Haziran 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 29 Aralık 2012.
- ^ Estabrook RW (December 2003). "A passion for P450s (remembrances of the early history of research on cytochrome P450)". Drug Metab. Dispos. 31 (12). ss. 1461-73. doi:10.1124/dmd.31.12.1461. PMID 14625342.
- ^ Estabrook RW, Cooper DY, Rosenthal O (1963). "The light reversible carbon monoxide inhibition of steroid C-21 hydroxylase system in adrenal cortex". Biochem Z. Cilt 338. ss. 741-55. PMID 14087340.
Konuyla ilgili yayınlar
- Parvez H, Reiss C (2001). Molecular Responses to Xenobiotics. Elsevier. ISBN 0-345-42277-5.
- Ioannides C (2001). Enzyme Systems That Metabolise Drugs and Other Xenobiotics. John Wiley and Sons. ISBN 0-471-89466-4.
- Richardson M (1996). Environmental Xenobiotics. Taylor & Francis Ltd. ISBN 0-7484-0399-X.
- Ioannides C (1996). Cytochromes P450: Metabolic and Toxicological Aspects. CRC Press Inc. ISBN 0-8493-9224-1.
- Awasthi YC (2006). Toxicology of Glutathionine S-transferses. CRC Press Inc. ISBN 0-8493-2983-3.